Original Article - DOI:10.33594/000000821
Accepted 30 September 2025 - Published
online 17 October 2025
Effect of Isobutylparaben and Phenylmercuric Acetate on Juvenile Female Rats with Special Reference to Development, Neurobehavior, and Histopathology
Keywords
Abstract
Background/Aims:
Isobutylparaben (IBP) and phenylmercuric acetate (PMA) are extensively applied in pharmaceuticals, cosmetics, and industrial chemicals, and are of concern for developmental toxicity. This study was conducted to compare and assess the subchronic effects of IBP and PMA on growth, neurobehavior, reproduction, and organ integrity in juvenile female rats, and establish no-observed-adverse-effect levels (NOAELs).Methods:
Juvenile female Sprague-Dawley rats (n=10/group) were given IBP (10, 20, 50 mg/kg/day) or PMA (2, 4, 8 mg/kg/day) by gavage for 70 days. Assays encompassed growth assessment, onset of puberty (vaginal opening), estrous cyclicity (vaginal smears), neurobehavioral examination (open-field activity, sensory reflexes, grip strength, motor activity), haematology, serum biochemistry, thyroid hormones (ELISA), organ weights, and histopathology of reproductive, hepatic, and renal tissues.Results:
IBP and PMA at high doses significantly inhibited terminal body weight, postponed vaginal opening, disrupted motor function, and affected exploratory behavior. Biochemical indicators revealed hepatic and renal stress and histological findings of hepatocellular hypertrophy and renal tubular degeneration. PMA was somewhat more toxic. NOAELs were 20 mg/kg/day (IBP) and 4 mg/kg/day (PMA).Conclusion:
Both IBP and PMA caused dose-dependent developmental toxicity, calling for more stringent pediatric exposure evaluations and more stringent chemical safety regulations. The results favor pediatric-specific risk assessment and more stringent monitoring of exposures during vulnerable growth stages.Introduction
Recently, there has been worldwide concern about the long-term health effects of environmental chemicals that are almost ubiquitous in our lives, especially endocrine-disrupting and neurotoxic chemicals [1]. In particular, the chemicals isobutyl paraben (IBP) and phenylmercuric acetate (PMA) received a great deal of attention because of their common use and ability for human exposure [2]. These chemicals can be commonly found in pharmaceuticals, cosmetics, food packaging, agricultural formulations, and in industrial work and applications. As a result of their biospheric persistence and bioactive nature, the ubiquitous use of these compounds in consumer products raises safety and health concerns, especially if exposure occurs during important periods of biological development [3, 4].
Isobutyl paraben is classified as an alkyl ester of p-hydroxybenzoic acid and is commonly used as a preservative in topical products, processed foods, and drugs [5]. Although considered safe at low concentrations, recent studies have suggested that parabens, such as IBP, have potential estrogenic activity and may interfere with endocrine systems [6]. IBP possesses a similar structure to estradiol and can bind to an estrogen receptor, influencing transcriptional pathways in ways that may affect development and reproduction [7]. Reports of reproductive defects, delayed puberty, and changes in hormone levels related to paraben exposure in animal models have been reported. Although these findings highlight potential risks, the toxicological evidence remains largely fragmented, particularly concerning long-term exposure during early developmental stages. The effects of sustained IBP exposure in juvenile female rats, spanning the critical period from weaning to sexual maturity, have to be thoroughly elucidated [8].
Phenylmercuric acetate, an organomercury compound, has historically been used as a fungicide, bactericide, and antiseptic [9]. Although its usage has declined due to regulatory interventions, its environmental persistence remains a significant concern. PMA and other organomercurials possess a well-established profile of toxicity, particularly to the central nervous system (CNS) and kidneys [10]. Mercury compounds exhibit a high affinity for sulfhydryl groups, disrupting enzymatic and cellular redox systems [11]. Developmental exposure to mercury, even at sublethal doses, has been associated with neurobehavioral impairments, motor dysfunction, and delayed reproductive maturation in both animals and humans [12]. The toxic effects of methylmercury are well documented; there exists a notable scarcity of systematic studies addressing the subchronic and developmental toxicity of PMA in juvenile organisms. This knowledge gap limits the ability to evaluate its risk potential during vulnerable life stages.
The juvenile period, spanning from weaning through sexual maturation, represents a critical window of vulnerability characterized by dynamic physiological, neurological, and hormonal changes [13]. During this time, organ systems undergo extensive growth, differentiation, and functional integration. Interference of such closely controlled processes by the exogenous toxicants may have long-term or irreversible health implications [14]. Adolescent animal model, especially Sprague-Dawley rats, provides a good platform to measure the above effects, since they have very close developmental milestones to those in human adolescents [15]. The ICH M3(R2) guideline that promotes the use of a subchronic study to determine the safety of the chemical in pediatric populations links to the importance of toxicological evaluation during this phase across international regulations [16]. Despite such awareness, there are also, to date few studies that have systematically explored the long-term outcomes of endocrine and heavy metal toxicants that are given at the juvenile stage in a controlled and dose-dependent way.
A thorough understanding of the toxicodynamic profiles for IBP and PMA during early life stages is especially needed. It seems plausible that these compounds are prevalent in both commercial and environmental settings, and exposure during growth and puberty is maintained. Regulatory decision-makers are currently relying on limited juvenile toxicity data to determine acceptable safety levels for these chemicals [17]. When looking at the extent literature, a comprehensive battery of endpoints (behavioral, neurofunctional, hematological, and histopathological) is not typically evaluated for long durations of exposure. This limits the ability to identify the levels of no-observed-adverse-effect levels (NOAEL) and weakens the underpinning for evidence-based regulations.
To fill the gaps, a well-structured 70-day subchronic toxicity study was performed in juvenile female Sprague-Dawley rats at postnatal age (PND) 23 to 26, an age that corresponds to early adolescence in humans and a very ideal window for evaluating the developmental and reproductive toxicity of environmental agents [18]. The compounds under investigation, IBP and PMA, were administered orally by gavage in three dose levels each: 10, 20, and 50 mg/kg/day for IBP, and 2, 4, and 8 mg/kg/day for PMA. Corn oil was used as the vehicle control. These dose ranges were selected based on previously reported subchronic tolerability profiles, pilot studies, and the need to identify threshold-dependent toxic effects. Daily administration was maintained for 70 consecutive days to cover key physiological transitions from prepuberty through sexual maturity.
To ensure a multidimensional assessment of toxicity, a comprehensive array of biological endpoints was employed. These included clinical observations, body weight and food consumption, functional observation battery (FOB) evaluations, sensory reactivity, hindlimb foot splay, grip strength, and motor activity assessments during the final week [19]. Developmental and reproductive progress was evaluated through estrous cyclicity tracking and the determination of age and weight at vaginal opening, a key indicator of puberty onset [20].
Blood samples were analyzed for hematological parameters such as hemoglobin, RBC, WBC, hematocrit, and reticulocyte count. Serum biochemistry was performed to assess liver and kidney function, including markers such as ALT, AST, creatinine, total protein, and electrolytes. Thyroid function was examined via ELISA assays for TSH and T4. At necropsy, major Organ weights and histological analyses were performed, with specific attention to the liver, spleen, thymus, adrenal glands, and reproductive tissues.
The study aims to examine and compare the subchronic impact of isobutylparaben and phenylmercuric acetate on growth, neurobehavioral parameters, reproductive development, and tissue histopathology in juvenile female rats after a 70-day oral exposure. The study aims to identify compound-specific patterns of systemic toxicity and establish potential no-observed-adverse-effect levels (NOAELs), providing essential data for the toxicological profiles of both compounds. The findings of the investigation aim to bridge critical knowledge gaps in juvenile toxicology and provide evidence-based insight for future regulatory assessments concerning developmental exposure to environmental chemicals.
Materials and Methods
Study Design and Animal Husbandry
This subchronic toxicity study (70-day) was performed by the ICH M3(R2) (2009) guidelines to ascertain the
effects of isobutylparaben (IBP) and phenylmercuric acetate (PMA) on growth, neurobehavior, development,
and
organ health in juvenile female rats. 80 Sprague-Dawley female rats aged 23-26 postnatal days (PND) and
weighing
40-55 g were randomly distributed into 8 distinct experimental groups, with each group comprising 10
individuals
(n = 10 per group).
Standard polypropylene cages with controlled environmental conditions (temperature = 22 ± 3°C; humidity =
55 ± 10%; 12:12 h light/dark cycle) were used to house the animals in pairs. Rats had ad libitum access to
a
certified rodent diet and filtered drinking water. Bedding was changed regularly, and health status was
monitored twice daily.
Dosing and Experimental Groups
Rats were administered IBP or PMA via oral gavage once daily for 70 consecutive days. IBP was tested at
10, 20,
and 50 mg/kg/day, and PMA at 2, 4, and 8 mg/kg/day. Corn oil was used as the vector for both compounds and
served as a control in the respective groups. The dosing amount was 10 mL/kg/day and was adjusted weekly
based
on individual body weights, as shown in Table 1.
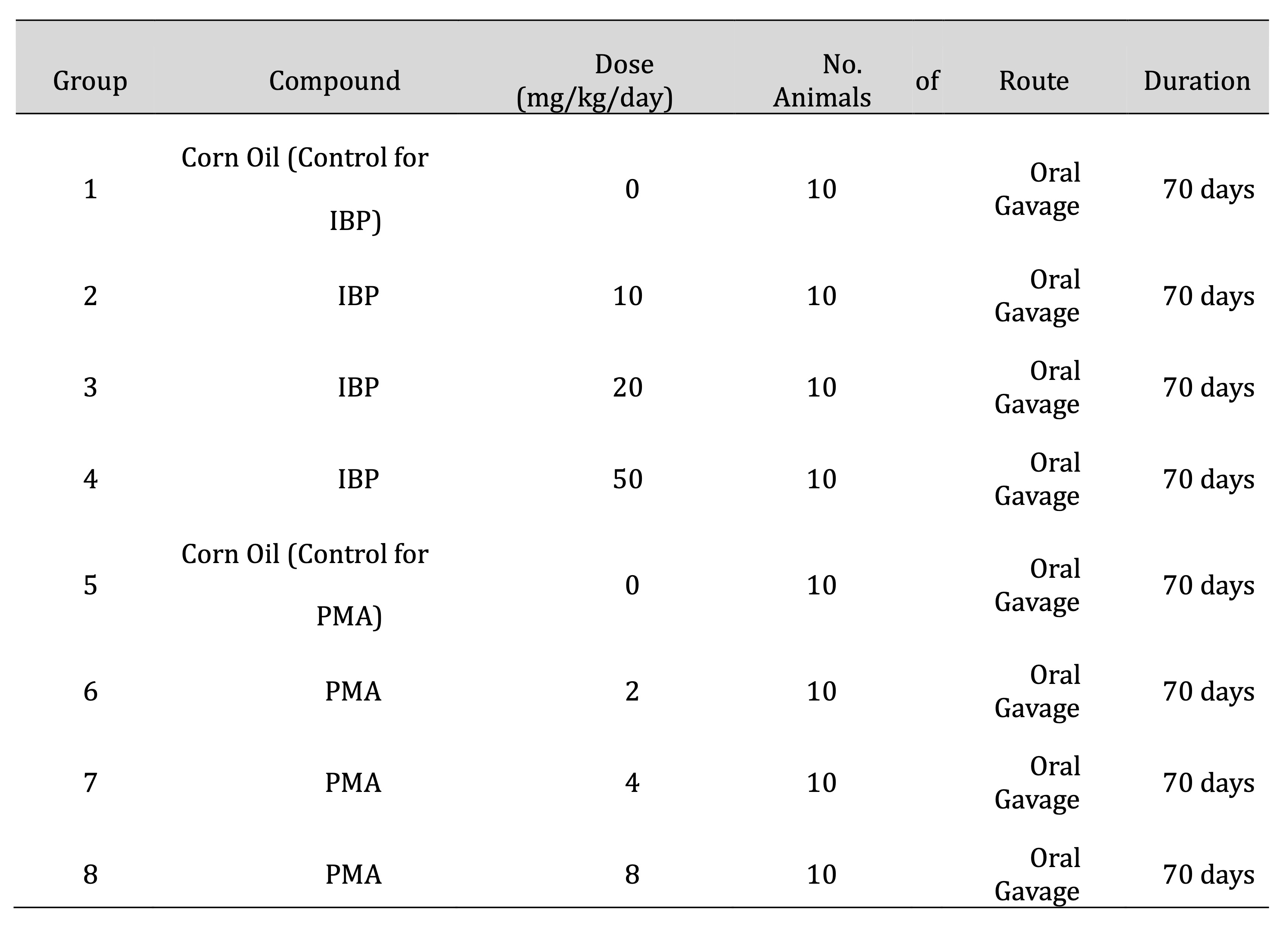
Table 1: Experimental grouping and dosing schedule for juvenile female rats exposed to isobutylparaben (IBP) or phenylmercuric acetate (PMA)
Clinical Observations and Body Weight Monitoring
Throughout the trial, every animal was checked twice a day for clinical symptoms and mortality. Body
weights
were measured on the first day before treatment and then every week until the trial was over. These
measurements
were used for monitoring growth and adjusting dosing volumes.
Food Consumption
Food intake was tracked every week by subtracting the residual feed from the amount provided at the start
of
each week. The data were averaged per cage and expressed as mean daily intake per animal.
Neurological Assessments (Functional Observation Battery)
Neurological evaluations were performed during the 10th week of the study and included:
Home Cage Observations
Rats were assessed for posture, unusual vocalizations, and convulsions without disturbing the cage.
Open Field Observations
Each rat was placed individually in a clean arena lined with absorbent paper and observed for 2 minutes.
Behavioral endpoints included gait, posture, arousal level, mobility, rearing, urination, defecation,
stereotypies, clonic/tonic movements, bizarre behavior, and vocalization.
Sensory Reactivity
Reflex responses were measured using standard stimuli, including approach, touch, tail pinch, click sound,
aerial righting, and pupillary response.
Hindlimb Landing Foot Splay
Once the heels were inked, the animals were dropped onto a recording sheet from a height of 30 cm. The
distance
between the hindlimb heel prints was measured from three trials, and the mean was used for analysis,
following
the method by Edwards and Parker [21].
Sexual Maturation Assessment
Beginning on Day 30, all animals were checked for vaginal opening every day. The age and corresponding
body
weight during vaginal opening were recorded to assess the onset of puberty.
Estrous cyclicity was assessed daily from Days 50–70 by vaginal smears collected with salinemoistened
swabs.
Smears were stained with methylene blue and examined microscopically (40×) to determine the stage of the
cycle:
proestrus (nucleated epithelial cells), estrus (anucleated cornified cells), metestrus (mixed cornified
cells
and leukocytes), and diestrus (predominantly leukocytes). Cycles of 4–5 days were considered regular,
whereas
prolonged diestrus or persistent estrus were classified as irregular.
Clinical Pathology Investigations
On Day 71, blood samples were obtained through retro-orbital puncture under isoflurane anesthesia for
clinical
pathology evaluation.
Hemoglobin, red blood cells (RBC), white blood cells (WBC), hematocrit (Hct), platelet count, mean
corpuscular
volume (MCV), mean corpuscular hemoglobin (MCH), mean corpuscular hemoglobin concentration (MCHC),
differential
leukocyte count (DLC), reticulocytes (Retic), and mean platelet volume (MPV).
Alanine aminotransferase (ALT), alkaline phosphatase (ALP), aspartate aminotransferase (AST), albumin,
blood
urea nitrogen (BUN), creatinine (Creat), gamma-glutamyl transferase (GGT), glucose, phosphate, potassium
(K⁺),
sodium (Na⁺), chloride, calcium (Ca²⁺), lactate dehydrogenase (LDH), total protein (T. Pro), total
bilirubin (T.
Bil), triglycerides (Trig), cholesterol (T. Chol), globulin (Glob), and albumin/globulin ratio (A/G).
Organ Weights
At necropsy on Day 71, animals were euthanized by CO₂ inhalation. The liver, kidneys, brain, ovaries, and
adrenal glands were excised and weighed. Weights were recorded to the nearest 0.01 g and expressed as
absolute
values.
Results
Clinical Observations and Mortality
No mortality occurred in any experimental group throughout the 70-day oral exposure period. All rats
appeared
generally healthy, with normal grooming, posture, and activity levels.
Transient signs of hypoactivity and piloerection were checked in a few groups from the IBP (50 mg/kg) and
PMA (8
mg/kg) groups during Weeks 4 to 6. These effects were mild and self-resolving.
Ophthalmological Evaluation
Ophthalmologic examinations performed pre-study and at termination using 1% tropicamide revealed no
abnormalities. Fundus, retina, lens, and ocular vasculature appeared normal in all groups. No signs of
cataract,
inflammation, or retinal degeneration were detected.
Body Weight and Food Consumption
All animals gained weight progressively across the 10-week study period. Both IBP- and PMA-treated rats
exhibited significantly lower body weight gain from Week 4 onwards compared to controls, with notable
differences at Weeks 6 and 9, as shown in Table 2. Average weekly food consumption was reduced transiently
in
treated groups during Weeks 4 to 6 but normalised by the end of the study, as mentioned in Table 3.
Fig. 1 illustrates the mean weekly body weight and daily food intake in the control, IBP (50 mg/kg), and
PMA (8
mg/kg) groups over a 10-week period. Treated groups showed reduced weight gain and transient decreases in
food
consumption, especially during Weeks 4–6.
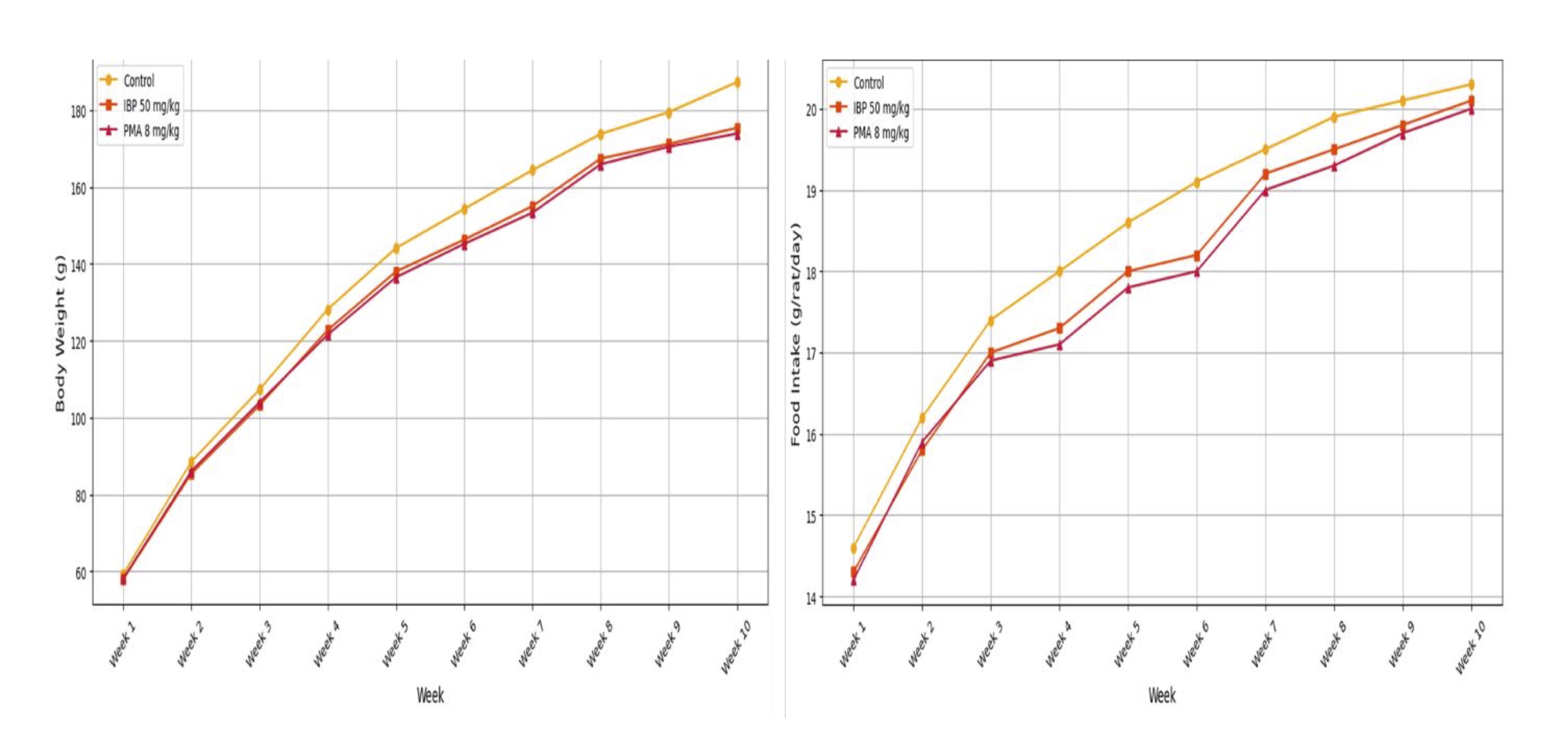
Fig. 1: Growth performance of juvenile female rats during 70-day oral exposure to isobutylparaben (IBP) or phenylmercuric acetate (PMA).
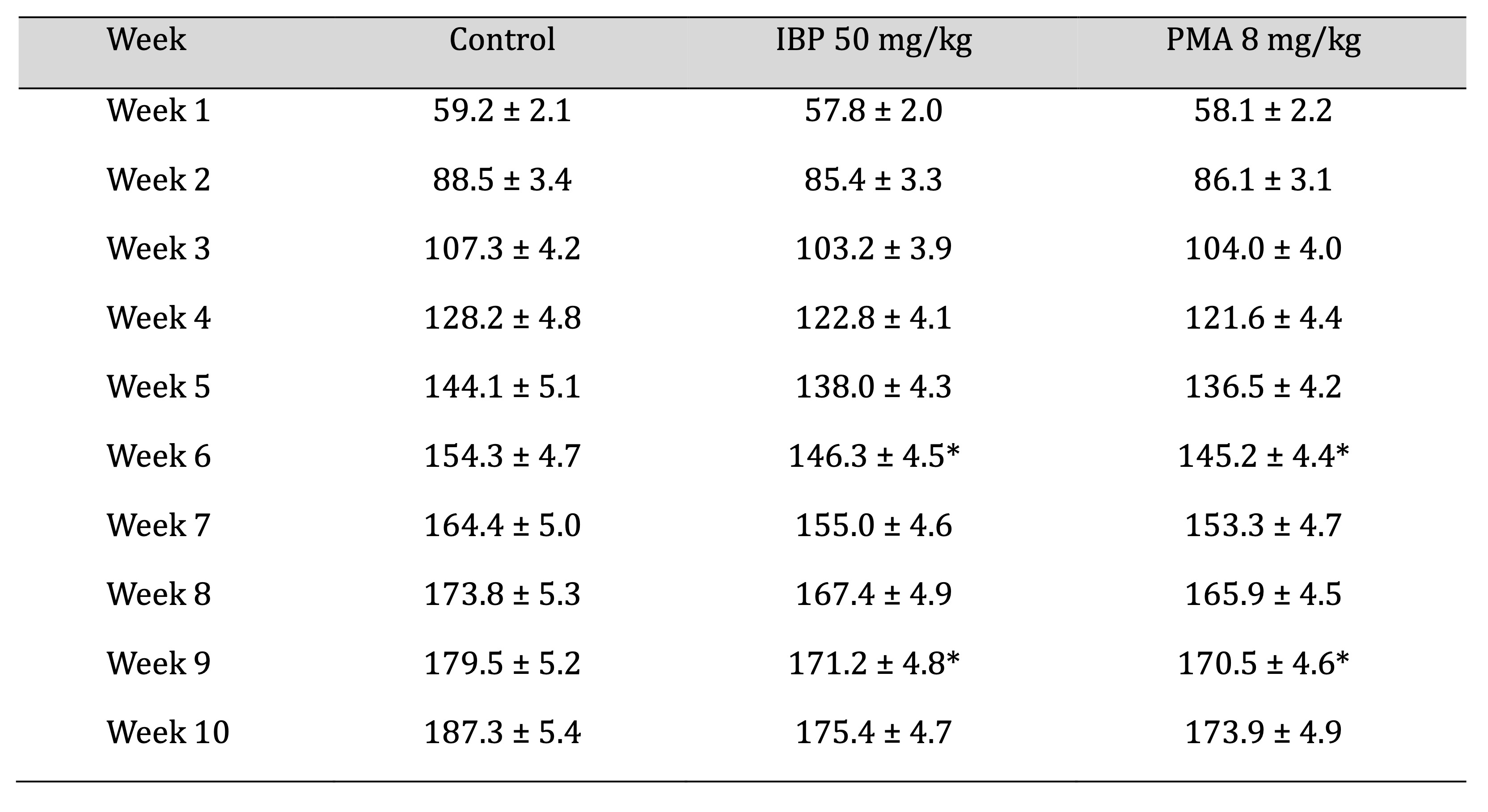
Table 2: Weekly mean body-weight progression of juvenile female rats during 70-day oral exposure to isobutylparaben (IBP) or phenylmercuric acetate (PMA)
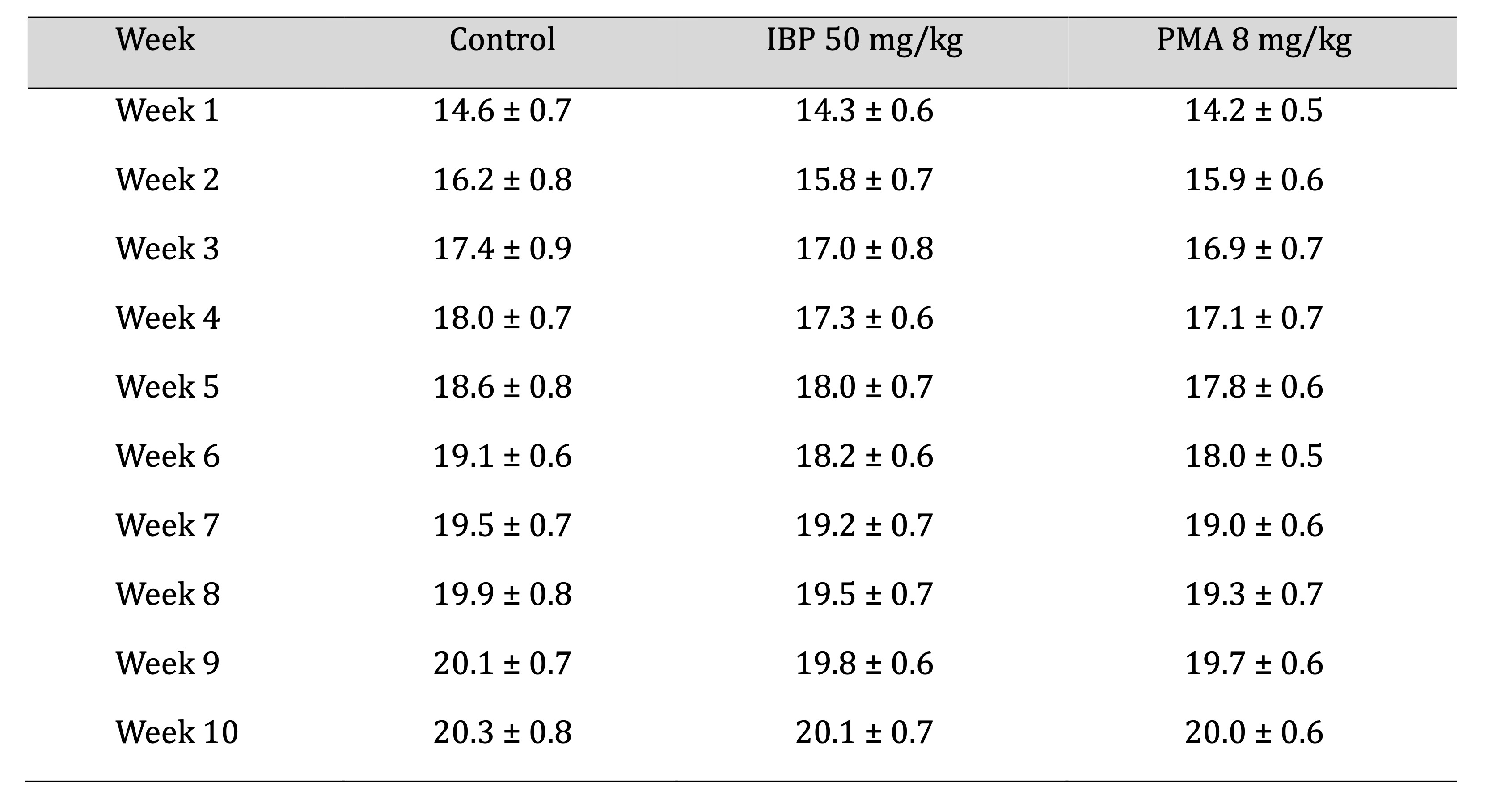
Table 3: Weekly mean food intake of juvenile female rats during 70-day oral exposure to isobutylparaben (IBP) or phenylmercuric acetate (PMA)
Functional Observational Battery and Reflex Integrity
Observations conducted in Week 10 revealed normal posture and absence of convulsions across all groups. In
open-field assessments, the number of rearing events per minute significantly declined in treated rats (p
<
0.05), suggesting a reduction in exploratory activity. Hindlimb foot splay distances were increased
dramatically
in both IBP and PMA groups compared to controls, indicative of neuromuscular impairment, as mentioned in
Table
4.
Fig. 2 illustrates the effects of isobutylparaben (IBP, 50 mg/kg/day) and phenylmercuric acetate (PMA,
8 mg/kg/day) on exploratory behavior and neuromuscular coordination in juvenile female rats after 70 days
of
exposure. Both compounds led to a reduction in rearing frequency (events/min), indicative of diminished
exploratory drive, while foot splay distance increased, reflecting impaired neuromuscular control.
Sensory reactivity parameters, including the click response, tail-pinch, and aerial righting reflexes,
remained
intact across all groups. Pupillary response to light was normal, and no abnormal vocalisations,
defecation, or
urination were observed during the testing period.
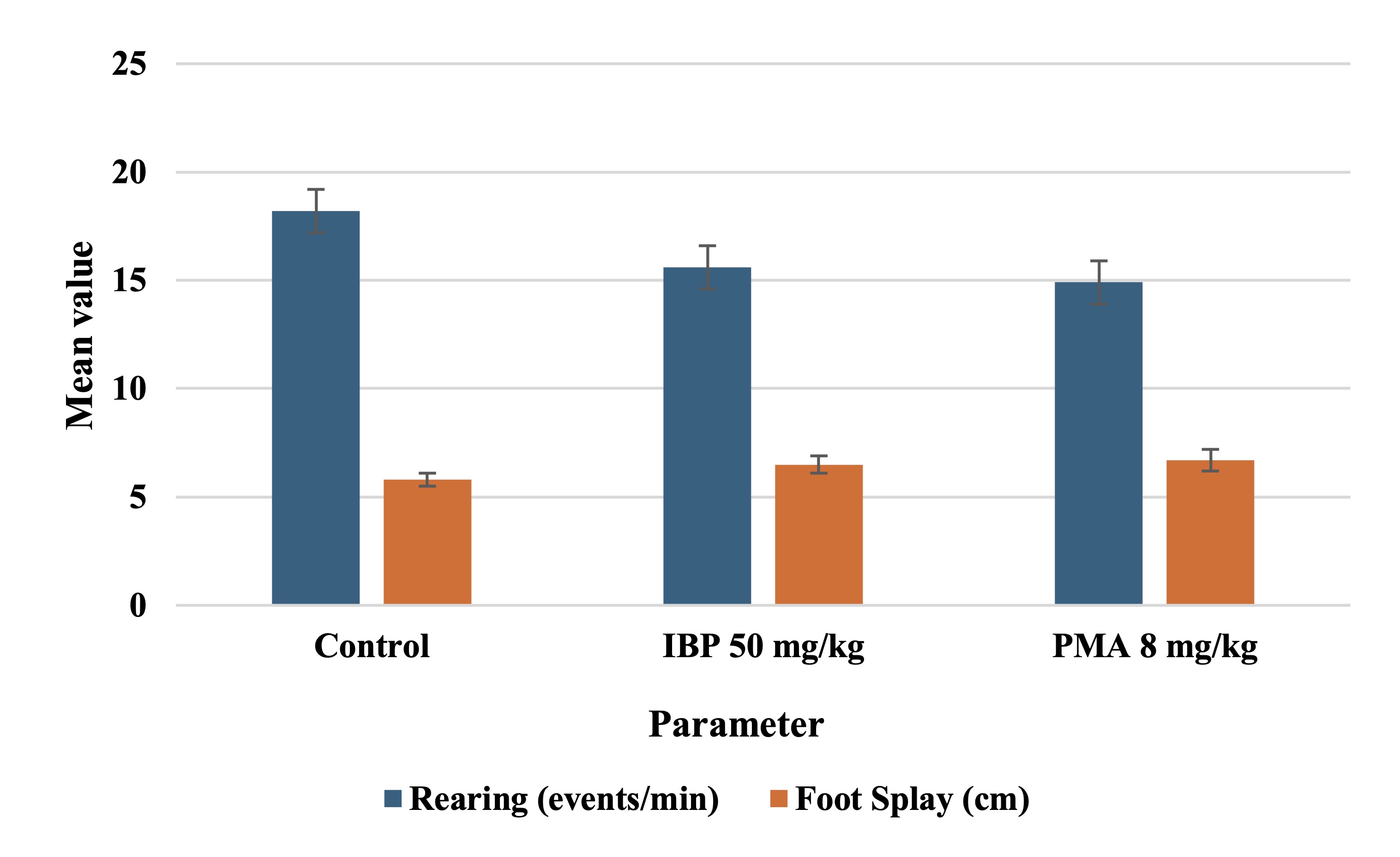
Fig. 2: Pubertal onset and body weight at vaginal opening in juvenile female rats exposed to isobutylparaben (IBP) or phenylmercuric acetate (PMA).
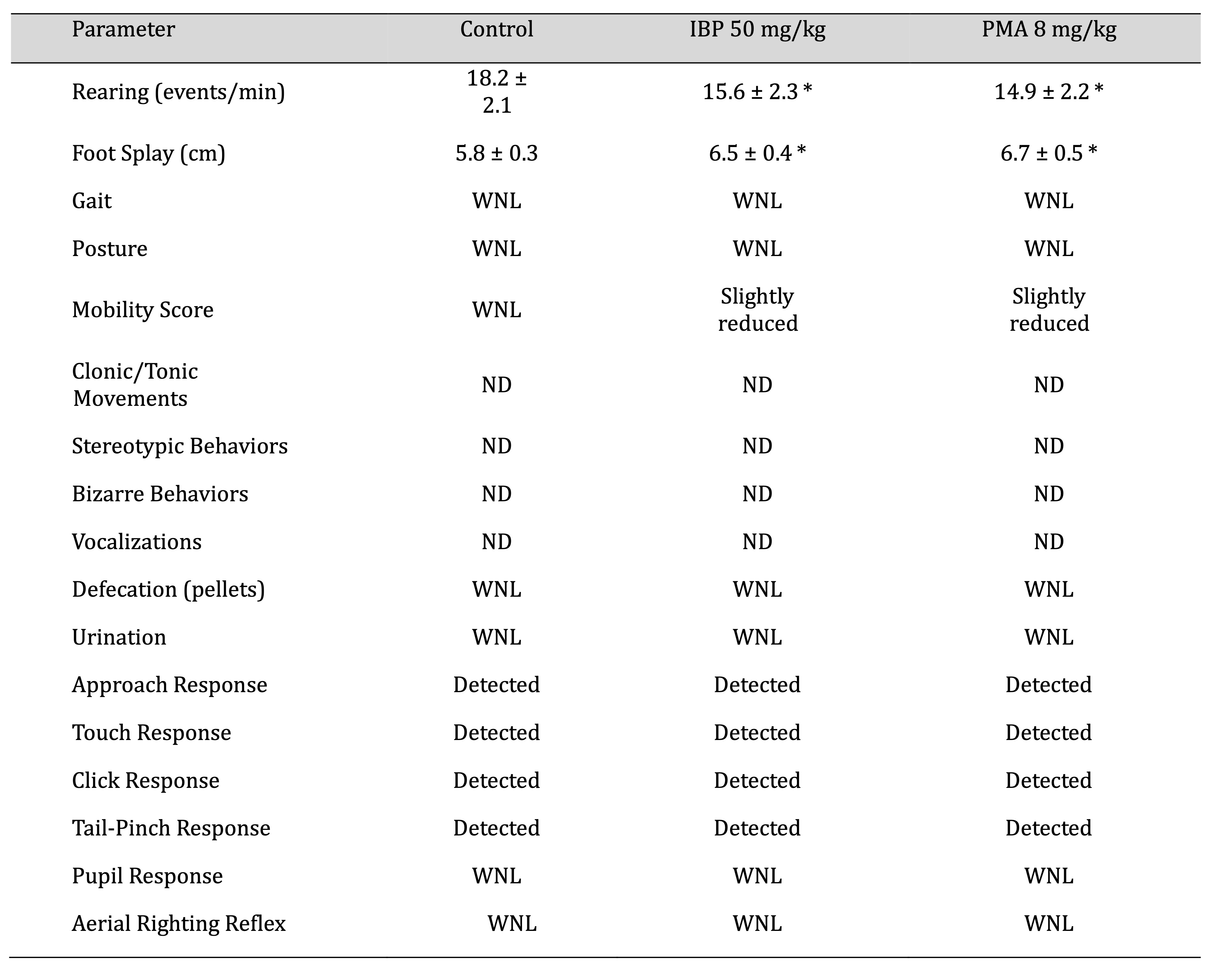
Table 4: Neurobehavioral assessment of juvenile female rats after 70-day oral exposure to isobutylparaben (IBP) or phenylmercuric acetate (PMA). Abbreviations WNL: Within Normal Range, ND: Not Detected
Sexual Maturation
Vaginal opening (VO), a key marker of pubertal onset, occurred significantly later (p < 0.05) in both
IBP-
and PMA-treated rats compared to controls. The mean body weight at VO was correspondingly higher in
treated
groups, indicating a delay in sexual maturation attributable to compound exposure, as shown in Table 5.

Table 5: Pubertal onset in juvenile female rats following 70-day oral exposure to isobutylparaben (IBP) or phenylmercuric acetate (PMA)
Estrous Cycle Monitoring
Estrous cyclicity was monitored in the final three weeks of exposure (Days 50–70). A significant
proportion of
high-dose IBP and PMA groups displayed irregular estrous cycles, characterised by extended diestrus or
disrupted
transitions between phases. The percentage of animals with regular 4–5 day cycles declined markedly in the
IBP
50 mg/kg and PMA 8 mg/kg groups (60% and 50%, respectively) compared to controls (90%). Prolonged diestrus
was
the most common abnormality, as shown in Table 6.
The mean cycle length was significantly extended in both high-dose groups (p < 0.05), as shown in Fig.
3,
further supporting endocrine disruption by these compounds.
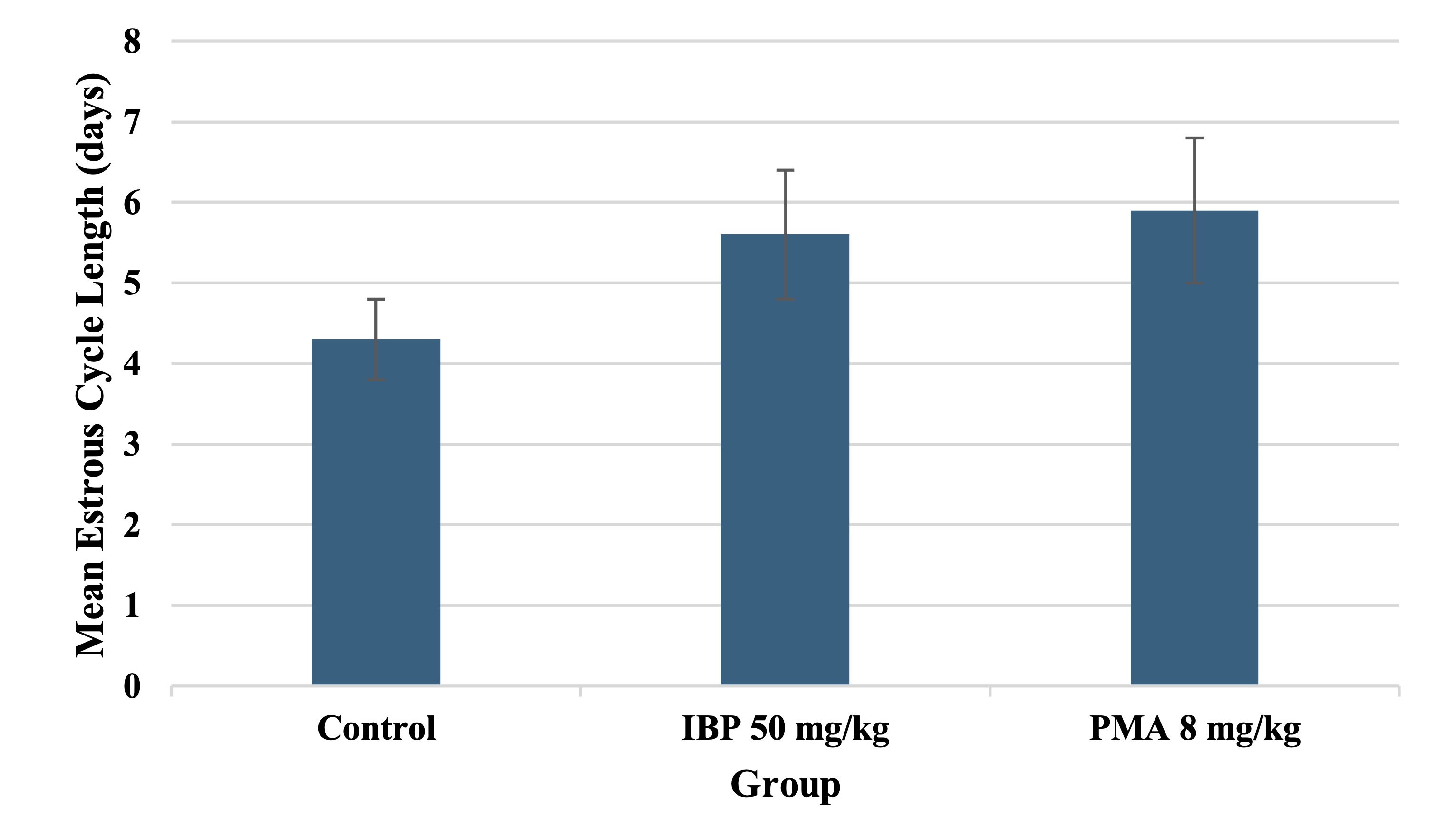
Fig. 3: Estrous cycle length in juvenile female rats following 70-day oral exposure to isobutylparaben (IBP) or phenylmercuric acetate (PMA).
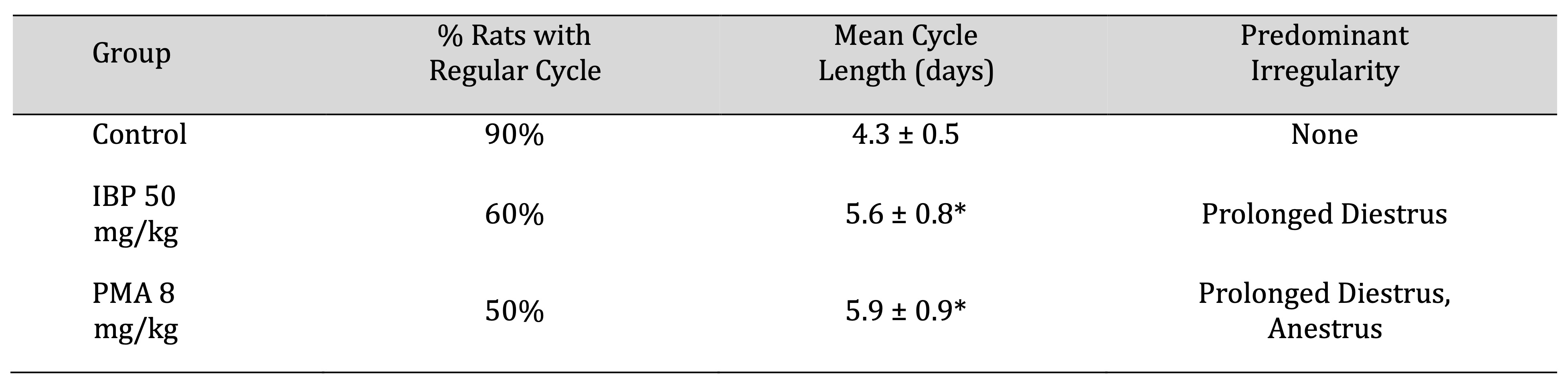
Table 6: Estrous-cycle characteristics of juvenile female rats after 70-day oral exposure to isobutylparaben (IBP) or phenylmercuric acetate (PMA)
Hematological Evaluation
Significant reductions in hemoglobin levels and red blood cell counts were observed in both treated groups
(p
< 0.05), indicative of mild anemia, as shown in Fig. 4. Other erythrocytic indices, such as hematocrit,
mean
corpuscular volume, and mean corpuscular hemoglobin concentration, showed marginal but non-significant
changes.
Platelet, white blood cell counts, and reticulocyte levels remained within physiological ranges,
suggesting
preserved immunological and thrombocytic function, as shown in Table 7.
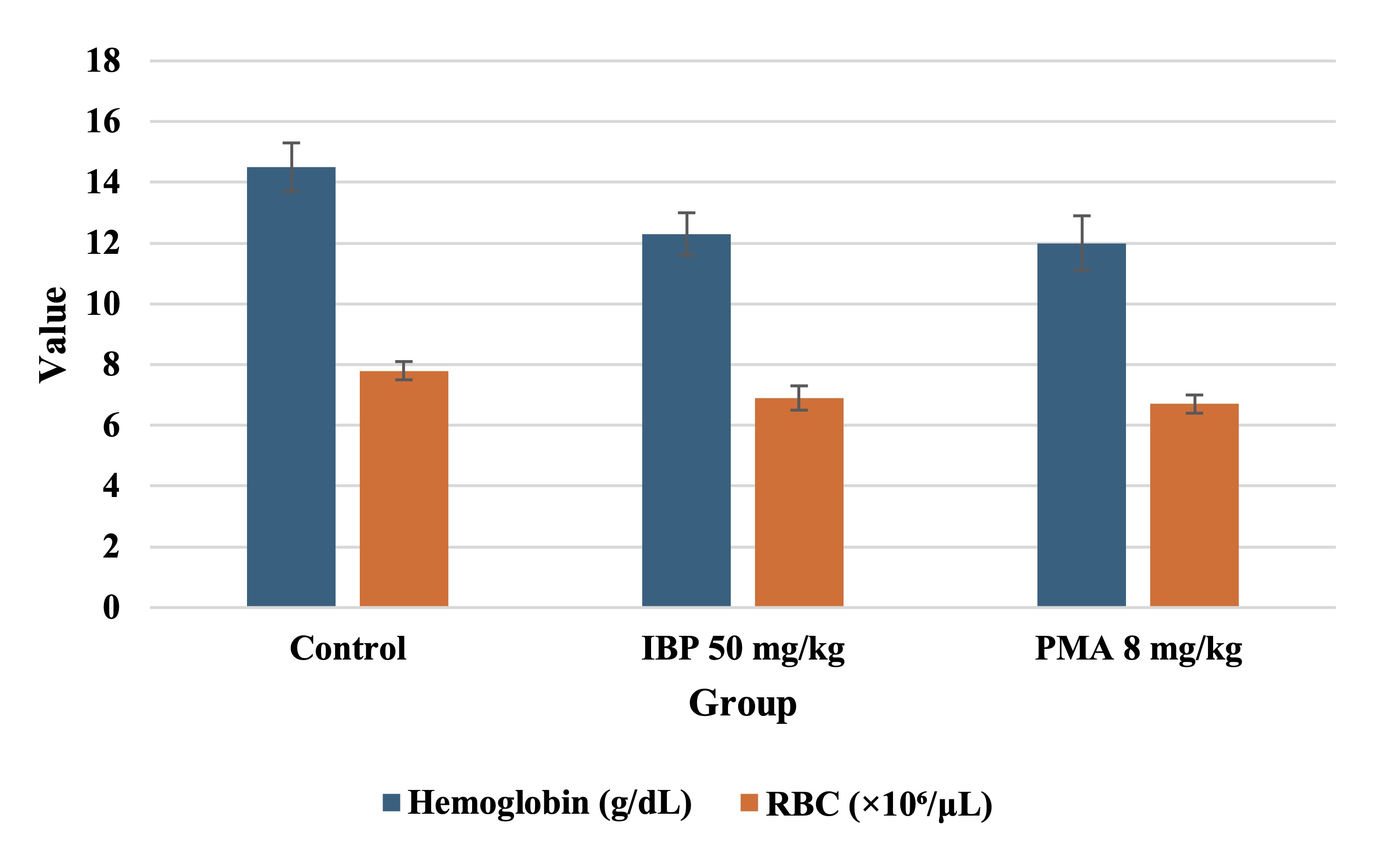
Fig. 4: Neurobehavioral performance of juvenile female rats after 70-day oral exposure to isobutylparaben (IBP) or phenylmercuric acetate (PMA).
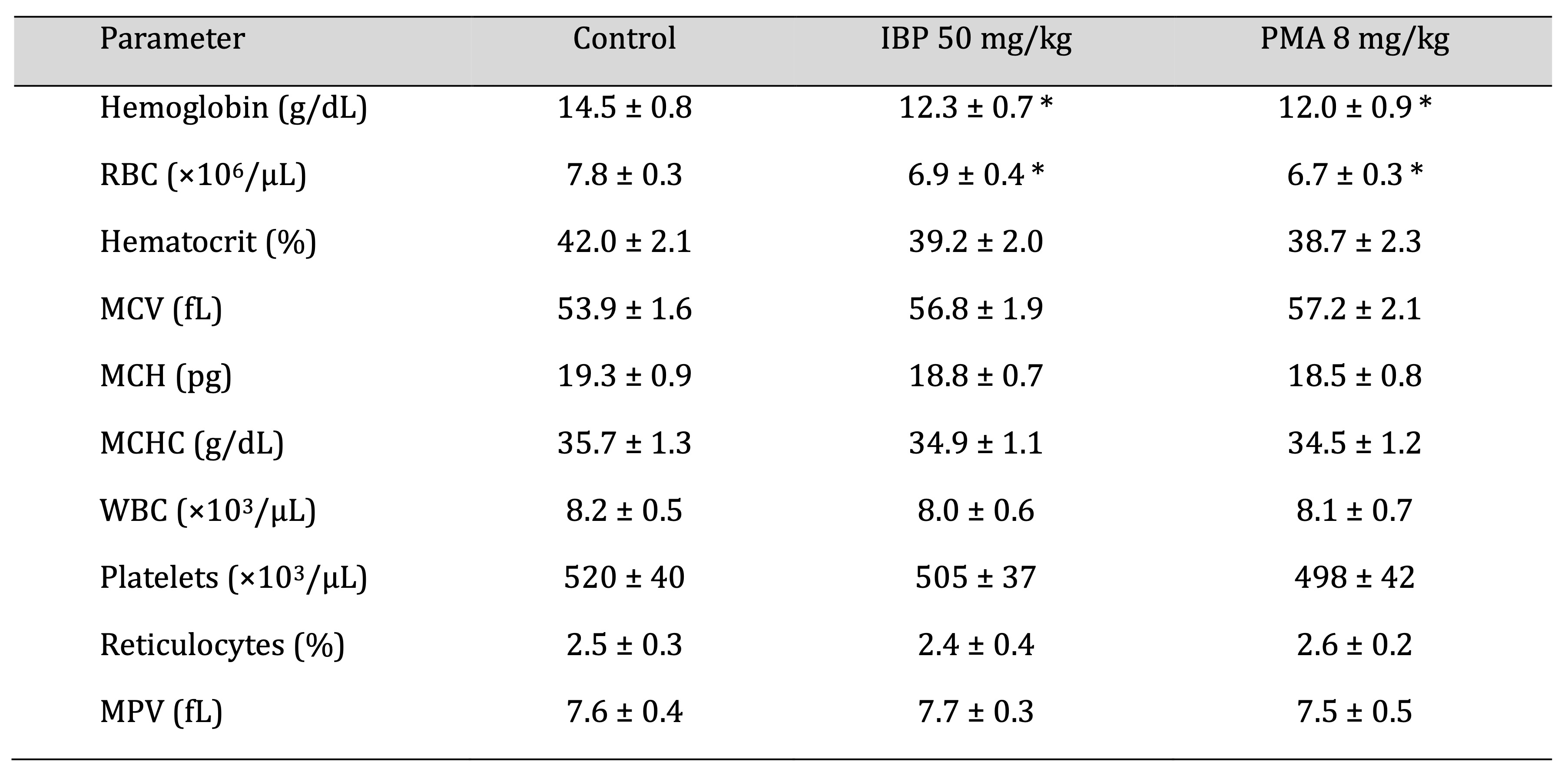
Table 7: Hematological parameters of juvenile female rats following 70-day oral exposure to isobutylparaben (IBP) or phenylmercuric acetate (PMA)
Fig. 4 illustrates the effects of isobutylparaben (IBP, 50 mg/kg/day) and phenylmercuric acetate (PMA, 8 mg/kg/day) on hemoglobin concentration and red blood cell (RBC) count in juvenile female rats after 70 days of oral exposure. Both treatment groups exhibited statistically significant reductions in hemoglobin and RBC levels compared to controls, indicative of mild anemia.
Clinical Biochemistry
Biochemical analysis revealed significantly elevated serum levels of alanine aminotransferase, aspartate
aminotransferase, creatinine, and blood urea nitrogen (BUN) in both IBP and PMA groups (p < 0.05),
indicating
hepatic and renal stress. Minor, non-significant shifts were observed in ALP, albumin, globulin, total
protein,
electrolytes (Na⁺, K⁺, Cl⁻), LDH, and glucose across groups. The A/G ratio remained stable, as shown in
Table 8.
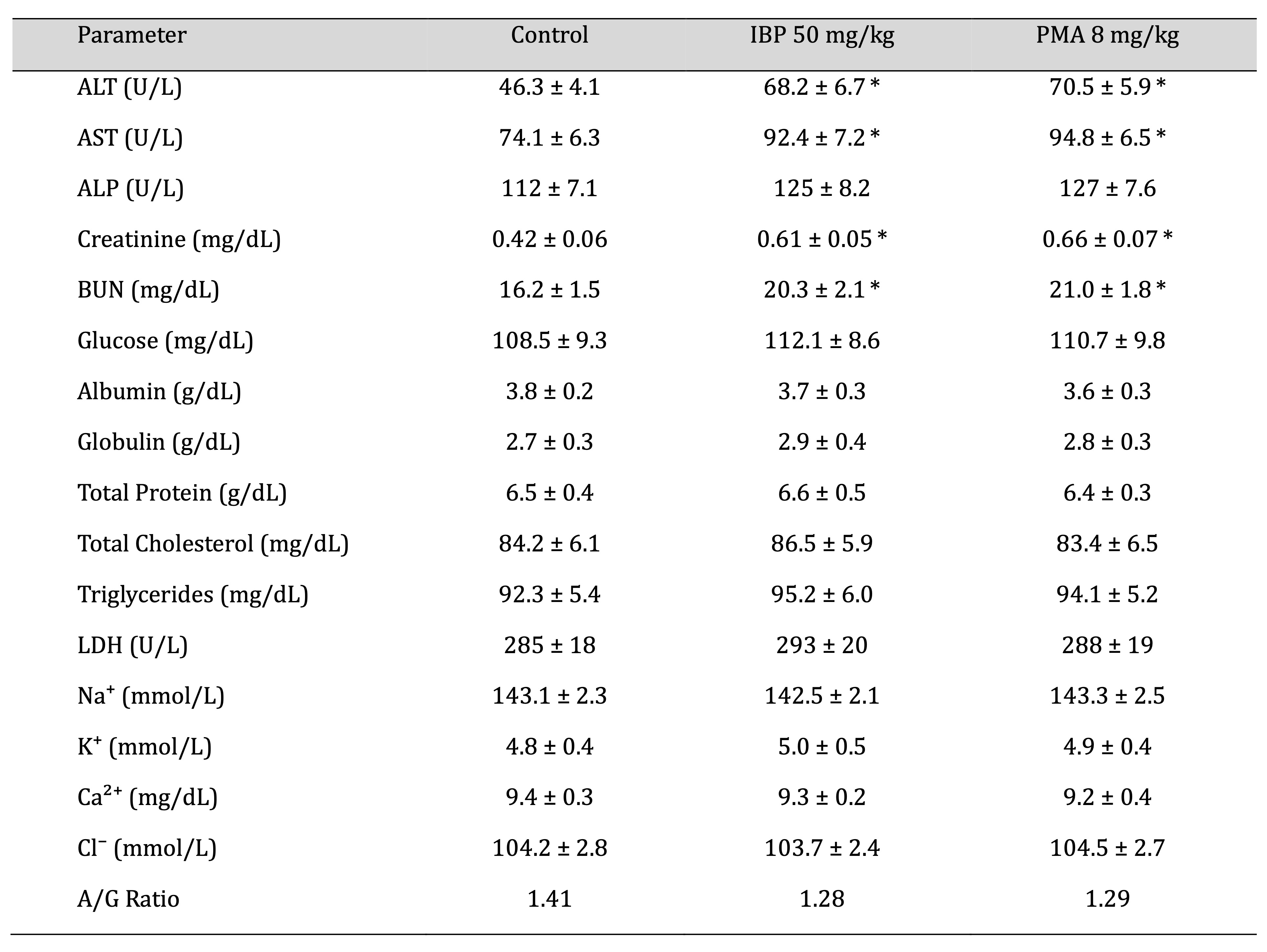
Table 8: Clinical biochemistry parameters of juvenile female rats after 70-day oral exposure to isobutylparaben (IBP) or phenylmercuric acetate (PMA)
Absolute Organ Weights
Both IBP and PMA groups exhibited significant increases (p < 0.05) in liver and kidney weights compared
to
controls, suggesting organ hypertrophy. Ovarian weights were slightly reduced, though not statistically
significant. Brain and adrenal weights showed no meaningful differences across groups, as shown in Table
9.
Fig. 5 depicts the absolute organ weights of the liver and kidneys in juvenile female rats after 70 days
of oral
exposure to isobutylparaben (IBP, 50 mg/kg/day) and phenylmercuric acetate (PMA, 8 mg/kg/day). Both
compounds
caused a statistically significant increase in liver and kidney weights compared to the control group,
suggesting organ hypertrophy and potential toxicological stress.
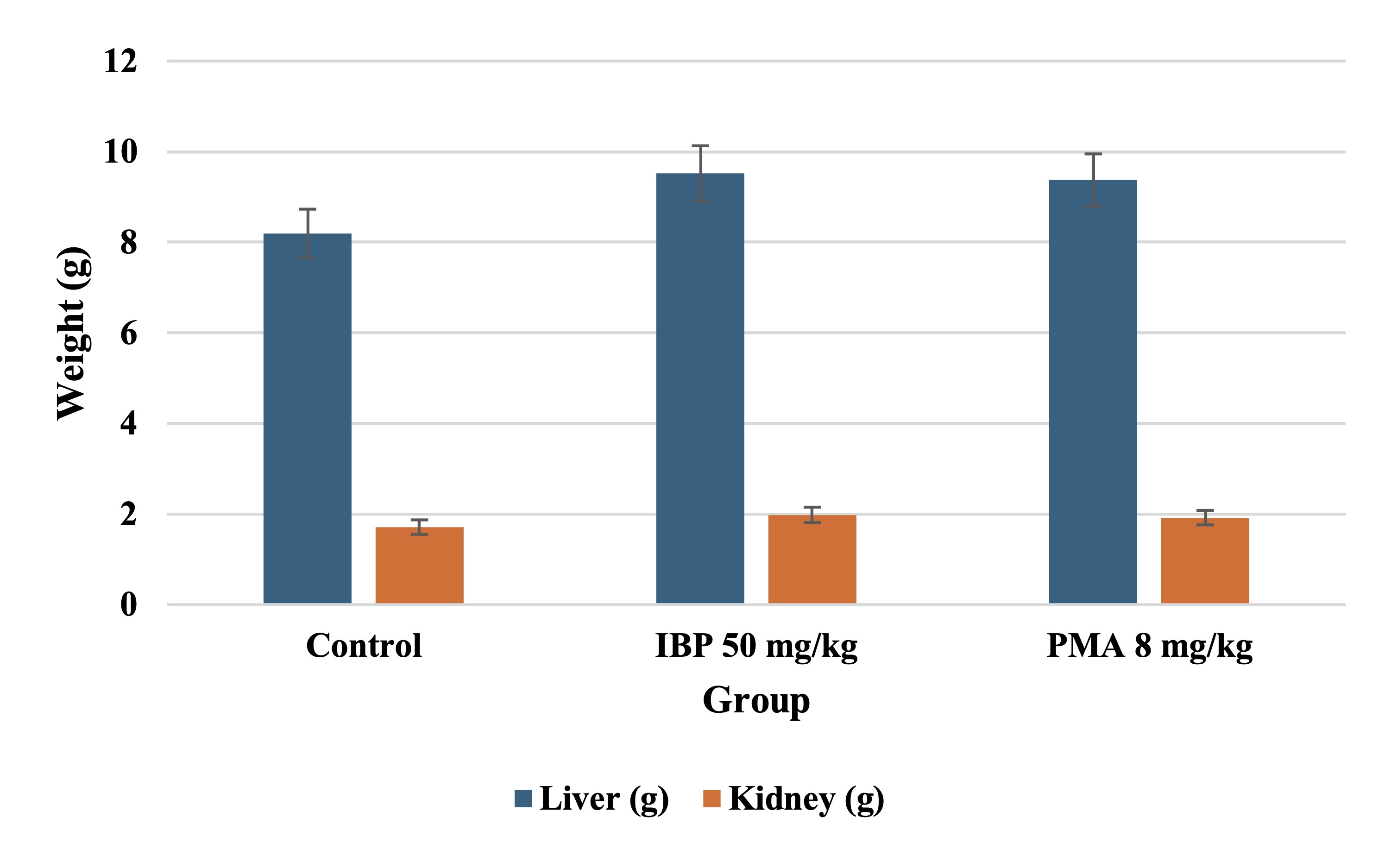
Fig. 5: Hematological changes in juvenile female rats after 70-day oral exposure to isobutylparaben (IBP) or phenylmercuric acetate (PMA).
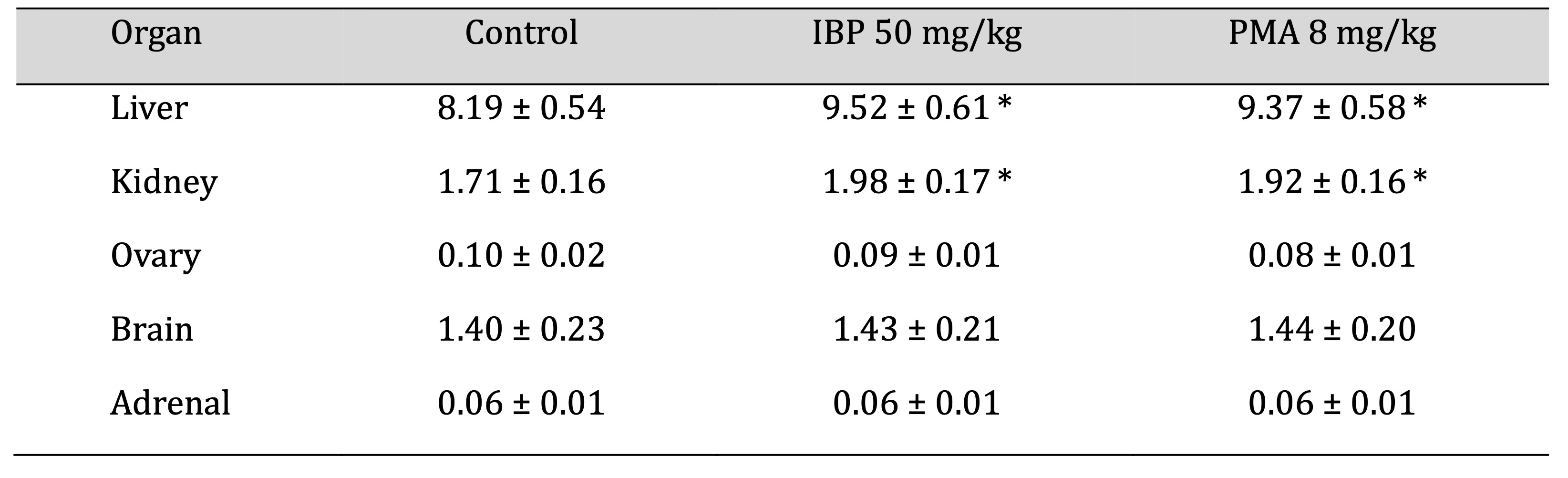
Table 9: Relative organ weights of juvenile female rats after 70-day oral exposure to isobutylparaben (IBP) or phenylmercuric acetate (PMA). Values are expressed as mean ± SD. Asterisks (*) indicate statistically significant differences compared to the control (p < 0.05)
Histopathological Findings
Microscopic examination revealed mild centrilobular hepatocellular hypertrophy and vacuolar degeneration
in
liver tissues from the treated groups. Kidneys displayed early signs of tubular dilation and epithelial
degeneration. Ovarian, adrenal, and brain tissues appeared histologically normal with no evidence of
necrosis,
inflammation, or structural disruption.
Discussion
This study systematically evaluated and compared the subchronic developmental and systemic toxicity profiles of isobutylparaben (IBP) and phenylmercuric acetate (PMA) in juvenile female rats over a 70-day oral exposure period, targeting a critical window of growth from weaning through sexual maturation. The data reveal distinct overlapping patterns of toxicological impact for both compounds, highlighting their potential to compromise physiological, neurological, and reproductive development in a dose-dependent manner.
Comparative analysis of the two agents underscores similarities in systemic toxicity while also delineating subtle differences in organ-specific and functional outcomes that may inform future risk assessment and regulatory decisions.
Both IBP and PMA exerted statistically significant reductions in body weight gain at their highest administered doses, with declines beginning around the fourth week of exposure and persisting until termination. Although the magnitude of reduction was comparable across compounds, PMA-treated animals exhibited slightly more pronounced suppression of terminal body weight and delayed recovery in food intake normalisation, suggesting a potentially greater disruption of metabolic processes or gastrointestinal function. This pattern aligns with PMA’s broader toxicodynamic reach, as mercury compounds are known to disrupt mitochondrial activity and nutrient absorption efficiency [22]. IBP-associated growth delays may be primarily endocrine-mediated, as supported by its estrogen receptor-binding affinity and reported interference with thyroid hormone regulation. These differential mechanisms warrant further mechanistic exploration to delineate compound-specific pathways affecting adolescent growth dynamics.
The onset of puberty, evaluated through weight and age at the vaginal opening, was significantly delayed in both treatment groups, with PMA causing a slightly longer postponement than IBP. This delay, alongside increased body weight at the time of VO, strongly implicates disruption of the hypothalamic-pituitary-gonadal axis by both agents. IBP, due to its structural similarity to endogenous estrogens, possibly exerts its effects through estrogen receptor modulation, resulting in feedback inhibition and subsequent delay in gonadotropin secretion. PMA, on the other hand, may act via indirect mechanisms, such as redox imbalance in neuroendocrine tissues or interference with zinc-dependent enzymes critical for reproductive hormone synthesis. Despite the overlapping phenotype, these divergent mechanistic underpinnings suggest that IBP and PMA disrupt pubertal maturation through different biochemical routes [23, 24].
Neurological assessments revealed compound-specific effects as well. While both IBP and PMA significantly reduced rearing behavior in open-field tests, indicative of suppressed exploratory drive, the foot splay test revealed a marginally larger increase in hindlimb splay distance in PMA-treated animals. This difference may reflect PMA’s neurotoxic potential via neuronal demyelination or synaptic transmission inhibition, consistent with existing literature on mercury-induced neurobehavioral toxicity. IBP's effects, though evident, appeared more confined to behavioral suppression than gross motor coordination deficits. The preservation of reflex integrity across all groups suggests that neither compound caused acute damage to sensory or spinal circuits; the subtle functional impairments underscore the need for more granular neurochemical and histological analyses in future studies.
From a hematological perspective, both compounds induced reductions in hemoglobin and red blood cell counts, pointing toward mild, non-regenerative anemia. The uniformity of this effect across compounds suggests a common impact on erythropoiesis or red cell lifespan, though PMA-treated animals exhibited slightly greater declines. Mercury’s known interference with heme synthesis and bone marrow function, this finding is consistent with previously documented organomercurial effects. IBP may indirectly suppress erythropoiesis through hepatic stress and systemic inflammation [25].
Clinical biochemistry data further support hepatotoxic and nephrotoxic trends, with both compounds significantly elevating ALT, AST, creatinine, and BUN levels. While the hepatic enzyme elevations were of similar magnitude between groups, PMA appeared to induce a slightly higher rise in creatinine, potentially indicating greater renal burden [26]. Histopathological observations corroborated these trends, with liver sections from both groups showing centrilobular hypertrophy and vacuolar degeneration, while kidney tissues exhibited early tubular epithelial damage. Ovarian and adrenal histology remained unaltered, suggesting that functional disturbances in these endocrine tissues might precede structural damage or fall below the detection threshold of routine microscopy. No histopathological changes were observed in the brain, although functional deficits identified through behavioral testing imply that neurotoxicity may occur without overt anatomical damage, at least at the light microscopy level.
Despite the strengths of the investigation, including the use of a well-characterised juvenile animal model, an extended exposure window, and a comprehensive battery of functional and biochemical endpoints, several limitations must be acknowledged. First, the study was conducted only in female rats, without hormonal assays such as serum estradiol, FSH, or LH, and without a post-exposure recovery phase, which limits detailed mechanistic understanding and long-term interpretation of the findings. The omission of these endocrine biomarkers is noteworthy because estradiol, FSH, and LH are key indicators of hypothalamic–pituitary– gonadal axis activity; previous work has demonstrated their value in linking chemical exposures to delayed puberty and disrupted reproductive development [27, 28]. The study exclusively utilized female rats, precluding the evaluation of sex-specific toxicological responses. Given that hormonal pathways and detoxification processes can differ markedly between sexes during adolescence, inclusion of male subjects would have supported the findings' generalizability. The absence of hormonal assays, such as serum estradiol, FSH, or LH, limited the mechanistic interpretation of pubertal delays and endocrine effects [29]. The 70-day study design, while sufficient to capture early maturation outcomes, did not include a post-exposure recovery phase, leaving open questions regarding the reversibility or persistence of observed toxic effects. Lastly, histopathological analyses, though insightful, lacked molecular validation such as immunohistochemistry or oxidative stress markers, which could have clarified underlying pathogenic processes [30]. Future studies should aim to address these limitations through multi-sex cohort designs, incorporation of hormonal and molecular biomarkers, and longitudinal follow-up to assess the permanence of developmental disruptions. Investigating the effects of these compounds in combination with other commonly encountered environmental agents may also provide ecologically relevant insights into mixture toxicity [31]. Exploration of gene expression changes, epigenetic modifications, and neurochemical imbalances will be instrumental in elucidating compound-specific mechanisms of action, particularly regarding neuroendocrine regulation and developmental plasticity.
The study provides compelling evidence that both isobutylparaben and phenylmercuric acetate exert dose-dependent toxic effects on juvenile female rats, with overlapping outcomes in growth retardation, delayed sexual maturation, mild neurobehavioral deficits, anaemia, and hepatic and renal stress. While both compounds induced similar toxicological patterns, PMA demonstrated slightly higher potency in several parameters, reflecting its broader biochemical reactivity [32].
The identification of distinct NOAELs for each compound (20 mg/kg/day for IBP and 4 mg/kg/day for PMA) offers valuable benchmarks for risk assessment in juvenile populations. These experimental thresholds are directly relevant to human health risk assessment because they align with early-life exposure scenarios where endocrine and neurodevelopmental processes are highly vulnerable. Given that both parabens and organomercurials are still detected in consumer products and environmental media, our findings highlight the need to reevaluate acceptable daily intake limits, particularly for infants, children, and adolescents. Regulatory agencies may consider incorporating adolescent-specific safety factors and endocrine-disruptor screening tests when updating guidelines for these chemicals. In addition, the evidence of subtle neurobehavioral changes without overt histopathological lesions underscores the importance of including sensitive neurodevelopmental endpoints in future human biomonitoring and policy frameworks.
Conclusion
Findings of this study highlight the developmental susceptibility of young organisms to two common environmental and pharmaceutical tolerants, isobutylparaben (IBP) and phenylmercuric acetate (PMA). These compounds resulted in dose-dependent, unequivocal, and unambiguous effects on the physiological growth, neurobehavioral functioning, reproductive maturation, and health endpoint of organs on prolonged oral exposure, carried out during the adolescent period. The puberty onset and exploratory behavior were severely delayed with high-dose treatments, and there were hematological effects, hepatotoxic and nephrotoxic biochemical dysfunctions, as well as histopathological liver and kidney damage. Although the patterns of systemic toxicity were quite similar in both compounds, PMA produced slightly stronger effects, which can be explained by neurotoxic and other biochemical activities of this compound. The identification of NOAELs for IBP and 4 mg/kg/day for PMA provides valuable benchmarks for refining pediatric safety thresholds. These findings highlight the importance of age-specific toxicological evaluations, as reliance on adult exposure data may fail to capture the unique susceptibilities of developing biological systems. The endocrinedisruptive and neuromodulatory profiles observed here support growing concerns about the silent, cumulative impact of low-dose, chronic exposures during early life stages. Enhanced scrutiny of chemical exposures during critical developmental windows is essential for informed regulatory decision-making. Future studies should extend these findings through longitudinal analyses, inclusion of male cohorts, mechanistic exploration via molecular markers, and combined exposure models that better reflect environmental realities. By advancing understanding of juvenile toxicodynamics, the current work contributes meaningfully to public health policy and the ongoing evolution of toxicological risk assessment frameworks.
Acknowledgements
The authors sincerely acknowledge Sharnbasva University for providing the research facilities and a supportive academic environment that enabled the successful completion of this work and We are deeply thankful to Dr. Somanath Reddy C Patil and Dr. Vijaykumar Malashetty his generous guidance and timely assistance that greatly contributed to the completion of this study.
Disclosure of AI Assistance
The authors declare that no AI tools or software were used in the preparation, writing, or editing of
this
manuscript
Statement of Ethics
The study was approved by Institutional Animal Ethics Committee (IAEC) from the Department of
Postgraduate
Studies and Research in Zoology, Sharnbasva University, Kalaburagi, Karnataka, India.
Disclosure Statement
The authors declare that they have no conflicts of interest.
References
| 1 | Thakur S, Chandra A, Kumar V, Bharti S. Environmental pollutants: endocrine
disruptors/pesticides/reactive dyes and inorganic toxic compounds, metals, radionuclides, and
metalloids and their impact on the ecosystem. In Biotechnology for environmental sustainability,
2025 Feb 11 (pp. 55-100). Singapore: Springer Nature Singapore.
https://doi.org/10.1007/978-981-97-7221-6_3 |
| 2 | Isaac RA, Subbarayalu R, Kumar MS, Martin TM, Lo SC, Santosh W. Human endocrine disruption: an
updated review of toxicological insights into parabens and phthalates. Toxicology and
Environmental
Health Sciences. 2025 May 29:1-4.
https://doi.org/10.1007/s13530-025-00264-w |
| 3 | Barabasz W, Pikulicka A, Wzorek Z, Nowak AK. Ecotoxicological aspects of the use of parabens in
the production of cosmetics. Technical Transactions. 2019 Dec 20;2019(Year 2019 (116)):99-124.
https://doi.org/10.4467/2353737XCT.19.126.11451 |
| 4 | Kim MJ, Kwack SJ, Lim SK, Kim YJ, Roh TH, Choi SM, Kim HS, Lee BM. Toxicological evaluation of
isopropylparaben and isobutylparaben mixture in Sprague- Dawley rats following 28 days of dermal
exposure. Regulatory Toxicology and Pharmacology. 2015 Nov 1;73(2):544-51.
https://doi.org/10.1016/j.yrtph.2015.08.005 |
| 5 | Safarova IR. Hydroxybenzoic acid derivatives and their biological activity. Processes of
Petrochemistry and Oil Refining. 2022;23(1).
|
| 6 | Liang J, Liu QS, Ren Z, Min K, Yang X, Hao F, Zhang Q, Liu Q, Zhou Q, Jiang G. Studying
paraben-induced estrogen receptor-and steroid hormone-related endocrine disruption effects via
multi-level approaches. Science of The Total Environment. 2023 Apr 15;869:161793.
https://doi.org/10.1016/j.scitotenv.2023.161793 |
| 7 | Amir S, Shah ST, Mamoulakis C, Docea AO, Kalantzi OI, Zachariou A, Calina D, Carvalho F,
Sofikitis
N, Makrigiannakis A, Tsatsakis A. Endocrine disruptors acting on estrogen and androgen pathways
cause reproductive disorders through multiple mechanisms: a review. International journal of
environmental research and public health. 2021 Feb;18(4):1464.
https://doi.org/10.3390/ijerph18041464 |
| 8 | Kuru BB, Kuru M. Reproductive behavior in female rats: Reproductive behavior in female rats.
Rats.
2023 Jul 13;1(1):21-6.
|
| 9 | Zhao M, Li Y, Wang Z. Mercury and mercury-containing preparations: history of use, clinical
applications, pharmacology, toxicology, and pharmacokinetics in traditional Chinese medicine.
Frontiers in Pharmacology. 2022 Mar 2;13:807807.
https://doi.org/10.3389/fphar.2022.807807 |
| 10 | Krout IN. The Role of Methylmercury Demethylation in Modulating Human Toxicokinetic Outcomes.
University of Rochester; 2023.
|
| 11 | Rupa SA, Patwary MA, Matin MM, Ghann WE, Uddin J, Kazi M. Interaction of mercury species with
proteins: towards a possible mechanism of mercurial toxicology. Toxicology research. 2023
Jun;12(3):355-68.
https://doi.org/10.1093/toxres/tfad039 |
| 12 | Azevedo LF, Karpova N, Rocha BA, Barbosa Junior F, Gobe GC, Hornos Carneiro MF. Evidence on
neurotoxicity after intrauterine and childhood exposure to organomercurials. International Journal
of Environmental Research and Public Health. 2023 Jan 7;20(2):1070.
https://doi.org/10.3390/ijerph20021070 |
| 13 | Sisk C, Lonstein JS, Gore AC. Critical periods during development: Hormonal influences on
neurobehavioral transitions across the life span. In Neuroscience in the 21st century 2013 (pp.
1715-1752). Springer, New York, NY.
https://doi.org/10.1007/978-1-4614-1997-6_61 |
| 14 | Grandjean P, Barouki R, Bellinger DC, Casteleyn L, Chadwick LH, Cordier S, Etzel RA, Gray KA, Ha
EH, Junien C, Karagas M. Life-long implications of developmental exposure to environmental
stressors: new perspectives. Endocrinology. 2015 Oct 1;156(10):3408-15.
https://doi.org/10.1210/en.2015-1350 |
| 15 | Vrolyk V, Tremblay C, Picut CA. Juvenile Toxicology. In Drug Discovery and Evaluation: Safety
and
Pharmacokinetic Assays 2024 Oct 22 (pp. 2337-2371). Cham: Springer International Publishing.
https://doi.org/10.1007/978-3-031-35529-5_121 |
| 16 | Srivastava AK, Negi G. Role of nonclinical programs in drug development. In The Quintessence of
Basic and Clinical Research and Scientific Publishing 2023 Oct 1 (pp.579-593). Singapore: Springer
Nature Singapore.
https://doi.org/10.1007/978-981-99-1284-1_35 |
| 17 | Sewell F, Lewis D, Mehta J, Terry C, Kimber I. Rethinking agrochemical safety assessment: A
perspective. Regulatory Toxicology and Pharmacology. 2021 Dec 1;127:105068.
https://doi.org/10.1016/j.yrtph.2021.105068 |
| 18 | Manservisi F. Reproductive and developmental toxicity study using Sprague-Dawley rats exposed
under various calendars to the weedkiller Glyphosate and commercial formulations Glyphosate-based.
2021.
|
| 19 | Malashetty V, Deshpande R, Patil S. Seventy‐Day Toxicity Study in Juvenile SpragueDawley Rats
with
Semicarbazide (SEM) from Weaning to Sexual Maturity. Journal of Toxicology. 2022;2022(1):5059761.
https://doi.org/10.1155/2022/5059761 |
| 20 | Sharma A. The role of puberty in shaping the ovarian reserve and reproductive lifespan (Doctoral
dissertation, University of Otago); 2024.
|
| 21 | Edwards PM, Parker VH. A simple, sensitive, and objective method for early assessment of
acrylamide neuropathy in rats. Toxicology and applied pharmacology.1977 Jun 1;40(3):589-91.
https://doi.org/10.1016/0041-008X(77)90083-7 |
| 22 | Han B, Lv Z, Han X, Li S, Han B, Yang Q, Wang X, Wu P, Li J, Deng N, Zhang Z. Harmful effects of
inorganic mercury exposure on kidney cells: mitochondrial dynamics disorder and excessive
oxidative
stress. Biological Trace Element Research. 2022 Apr;200(4):1591-7.
https://doi.org/10.1007/s12011-021-02766-3 |
| 23 | Rivera-Núñez Z, Kinkade CW, Zhang Y, Rockson A, Bandera EV, Llanos AA, Barrett ES. Phenols,
parabens, phthalates and puberty: a systematic review of synthetic chemicals commonly found in
personal care products and girls' pubertal development. Current environmental health reports. 2022
Dec;9(4):517-34.
https://doi.org/10.1007/s40572-022-00366-4 |
| 24 | Odenbro A. Effects on reproduction and hormone metabolism. In Biological Effects of Organolead
Compounds 2020 Mar 25 (pp. 161-175). CRC Press.
https://doi.org/10.1201/9780429282980-13 |
| 25 | Akmal H, Ahmad S, Abbasi MH, Jabeen F, Shahzad K. A study on assessing the toxic effects of
ethyl
paraben on rohu (Labeo rohita) using different biomarkers; hematobiochemical assays, histology,
oxidant and antioxidant activity, and genotoxicity. PLOS ONE. 2024 May 6;19(5):e0302691.
https://doi.org/10.1371/journal.pone.0302691 |
| 26 | Nicholson JK, Timbrell JA, Sadler PJ. Proton NMR spectra of urine as indicators of renal damage.
Mercury-induced nephrotoxicity in rats. Molecular Pharmacology. 1985 Jun 1;27(6):644-51.
https://doi.org/10.1016/S0026-895X(25)12559-5 |
| 27 | Parent AS, Teilmann G, Juul A, Skakkebaek NE, Toppari J, Bourguignon JP. The timing of normal
puberty and the age limits of sexual precocity: variations around the world, secular trends, and
changes after migration. Endocrine Reviews. 2003 Oct;24(5):668693.
https://doi.org/10.1210/er.2002-0019 |
| 28 | Ye X, Pan W, Zhao S, Zhao Y, Zhu Y, Liu J, Liu W. Relationships of pyrethroid exposure with
gonadotropin levels and pubertal development in Chinese boys. Environmental Science & Technology.
2017 Jun;51(11):6379-6386.
https://doi.org/10.1021/acs.est.6b05984 |
| 29 | Yesumanipreethi S, Nirmal Magadalenal N, Moses Inbaraj R. Impact of phthalates and parabens on
the
neurobehavioral and reproductive function: a review. InProceedings of the Zoological Society 2021
Dec (Vol. 74, No. 4, pp. 572-590). New Delhi: Springer India.
https://doi.org/10.1007/s12595-021-00408-z |
| 30 | Lee JD, Bae JS, Kim HY, Song SW, Kim JC, Lee BM, Kim KB. Repeated-dose toxicity and
toxicokinetic
study of isobutylparaben in rats subcutaneously treated for 13 weeks. Archives of Toxicology. 2024
Jul;98(7):2231-46.
https://doi.org/10.1007/s00204-024-03741-2 |
| 31 | Panghal A, Jena G. Single versus intermittent cycle exposure effect of 6mercaptopurine in
juvenile
Sprague-Dawley rat: a germ cell-specific mechanistic study. Naunyn-Schmiedeberg's Archives of
Pharmacology. 2024 May;397(5):3155-68.
https://doi.org/10.1007/s00210-023-02797-8 |
| 32 | Ullah H, Khan MF, Hashmat F. In vitro, concentration and time-dependent effect of phenyl
mercuric
acetate on the chemical status of glutathione (GSH). Gomal University Journal of Research. 2012
Dec
31;28(2):1-8.
|