Effect of Lychee Peel and Seed Flour Consumption on the Anti/Pro-Oxidant System and Cardiomyocyte Contractile Function
bPostgraduate Program in Physiological Sciences, Center of Health Sciences, Federal University of Espírito Santo, Vitória, Espírito Santo, Brazil,
cCenter of Health Sciences, Department of Pharmaceutical Sciences, Federal University of Espírito Santo, Vitória, Espírito Santo, Brazil
Keywords
Abstract
Background/Aims:
Type 2 diabetes mellitus (T2DM) represents a high risk for developing cardiovascular diseases, with alterations in contractile function and calcium (Ca2+) handling. In addition, there is an increase of reactive oxygen species in T2DM, with the pathways of altered glucose metabolism, oxidative damage to pancreatic β-cells, and endothelial dysfunction being involved in this process. Studies have shown that both the extract and lychee peel and seed flour are rich in antioxidant phenolic compounds, which could be beneficial in preventing and/or reversing oxidative stress (OS) in obesity associated with type 2 diabetes mellitus (T2DMOb). However, the relationship between oxidative stress in T2DMOb and the involvement of lychee peel and seed flour is still not well understood. This study aimed to evaluate the effect of lychee peel and seed flour consumption on the anti/pro-oxidant system and cardiomyocyte contractile function in obese rats induced to T2DM.Methods:
Obesity in Wistar rats (n = 38) was induced by a high-fat diet and, later, they were induced to T2DM. The experimental protocol consisted of a total period of 17 weeks and was divided into four moments (Figure 1): 1) obesity induction (4 weeks); 2) maintenance of obesity (8 weeks); 3) induction of T2DM (12th week) in obese (Ob) rats and redistribution of groups; and 4) obesity and T2DM maintenance and treatments with lychee peel and seed flours (5 weeks). After 12 weeks, the Ob rats were randomized into T2DMOb (n = 8), T2DMOb treated with lychee peel flour (T2DMObPF, n = 10), and T2DMOb treated with lychee seed flour (T2DMObSF, n = 10). Analyzes of the nutritional and metabolic profiles, cardiac remodeling, and OS biomarkers were evaluated. Contractile function by isolated cardiomyocyte analysis and Ca2+ handling was determined.Results:
Treatments with lychee peel and seed flour were not able to change body weight, adiposity, biochemical and cardiac morphological parameters, or OS biomarkers in relation to T2DMOb animals. Lychee treatments did not accentuate the elevation of fractional shortening visualized in T2DMOb. Regarding relaxation, the maximum rate of relaxation was higher in the T2DMOb group compared to the Ob group, but the lychee treatments did not promote positive alterations in this parameter. In addition, the time to 50% relaxation was also longer in the presence of T2DM (T2DMOb > Ob); the treatments with lychee peel flour favored a reduction in the time to 50% relaxation. T2DMOb rats presented an increase in diastolic Ca2+ in relation to the Ob group; the treatment with lychee seed flour reduced this parameter, despite not improving the time to 50% decay Ca2+ and, consequently, relaxation. Furthermore, the treatments with lychee peel and seed flour did not change the sensitivity of myofilaments to Ca2+ in T2DMOb animals.Conclusion:
The treatments with lychee peel and seed flour showed no significant effects on anti/pro-oxidant parameters. However, lychee peel flour demonstrated notable effectiveness in reducing myocardial relaxation time in a T2DMOb model. These results suggest that while the antioxidant properties of lychee seed and peel flours were not significantly altered, lychee peel flour may have specific therapeutic potential to improve cardiac function in T2DMOb.Introduction
Diabetic cardiomyopathy, characterized by alterations in the structure and function of the myocardium unrelated to vascular pathologies, is a condition that can be found in individuals with type 2 diabetes mellitus (T2DM) [1]. This disease occurs when the body does not use the insulin produced by the pancreas or it is not able to produce insulin to control the blood glucose level [2]. Insulin regulates glucose in the blood and ensures energy for the body.
Evidence indicates a positive influence between central obesity, T2DM, and increased risk of cardiac remodeling, characterized by left ventricular hypertrophy [3]. Obesity and hyperglycemia associated with T2DM cause oxidative stress due to the increased release of reactive oxygen species (ROS) [4]. Free radicals in excess can cause biomolecular damage that compromises health; however, they can be fought by antioxidants produced by the body or absorbed by food [5].
According to the World Health Organization [2], T2DM can be controlled with physical activity and dietary planning. It has been described that food sources of bioactive compounds, such as those rich in phenolic compounds with high antioxidant capacity, can be beneficial in preventing and/or reversing oxidative stress [6].
Lychee (Litchi chinensis Sonn. ) is a tropical to subtropical fruit that originates in Southeast Asia and belongs to the Sapindacea family. It is popular for its attractive color and flavor, and its by-products, such as its peel and seed, are generally discarded [7]. However, these residuals can be used as an important alternative for accessing foods rich in nutrients and bioactive compounds [8]. Some studies have shown that both the lychee seed extract and the flours from the peel and seed of this fruit are rich in phenolic compounds [9, 10].
In this context, research has highlighted the beneficial effects of lychee in T2DM [11, 12]. Man et al [11]. studied the effect of lychee seed extract in rats induced to diabetes and found a reduction in blood glucose levels compared to treated diabetic animals. The authors also showed that the flavonoids present in lychee seed extract were able to delay glucose release from carbohydrate consumption, suppressing postprandial hyperglycemia [13]. Thus, these findings show the antioxidant capacity of phenolic compounds. Another study carried out by LV et al [4]. highlights a positive aspect between the higher content of total flavonoids and greater hypoglycemic potential.
Despite the beneficial effects related to lychee, most studies have used pulp, seed, and flower extracts [4, 10]. Thus, the great potential of lychee may be related to the direct use of the peel and seed flours of this functional food in the preparation of breads, cakes, and cookies or in their addition to foods with the aim of improving their nutritional and energetic constitution, as well as lychee being a source of bioactive substances [14]. In addition, lychee peel and seed flours can be good alternatives to bioactive compounds in promoting health benefits, specifically to the cardiovascular system.
Considering the possible beneficial role of lychee and the lack of studies and information about the effects of consumption of lychee peel and seed flours on T2DMOb, the current study aimed to evaluate the role of lychee peel and seed flours on T2DMOb rats, specifically evaluating their effects on anti/pro-oxidant systems and cardiomyocyte contractile function. Our hypothesis was that these lychee treatments would attenuate the oxidative damage and contractile impairment caused by T2DMob.
Materials and Methods
Animal Care
Thirty-day-old male Wistar rats (≅ 150g) obtained from the Animal Center of the Federal University of Espírito Santo (Vitória, Espírito Santo, Brazil) were individually caged. The animals were kept under controlled temperature conditions (24 ± 2°C), relative humidity (60% ± 5%), and inverted 12-hour lighting cycles. Experimental procedures were performed in accordance with the Brazilian National Ethics Council (CONEP) and approved by the Ethics Committee of the Federal University of Espírito Santo (CEUA-UFES 24/2017).
Experimental Protocol
After a period of 7 days for acclimatization, the rats were randomized into two groups: standard diet (SD, n = 10) and saturated high-fat diet (HFD, n = 38). The SD group was fed with a standard diet (Nuvilab CR1-Nuvital, Colombo, Paraná, Brazil) containing 13.9% of its kcal from fat, 55.9% from carbohydrates, and 30.2% from protein. The HFD animals were fed a saturated high-fat diet (Nutriave Alimentos®, Vitória, Espírito Santo, Brazil) containing 37.6% of its kcal from fat, 44.6% from carbohydrates, and 17.8% from protein) [15]. All animals had free access to water and chow (40g/day).
During the experimental protocol, body weight was recorded weekly, and nutritional parameters such as food consumption (FC), caloric intake (CI), and feed efficiency (FE) were measured daily. CI was calculated by the mean FC x dietary energetic density. FE, the ability to turn consumed calories into body weight, was determined by the following formula: mean body weight gain (g)/total CI (kcal).
The experimental protocol consisted of a total period of 17 weeks and was divided into four moments (Fig. 1): 1) obesity induction (4 weeks); 2) maintenance of obesity (8 weeks); 3) induction of T2DM (12th week) in obese rats and redistribution of groups and; 4) obesity and T2DM maintenance and treatments with lychee peel and seed flours (5 weeks).
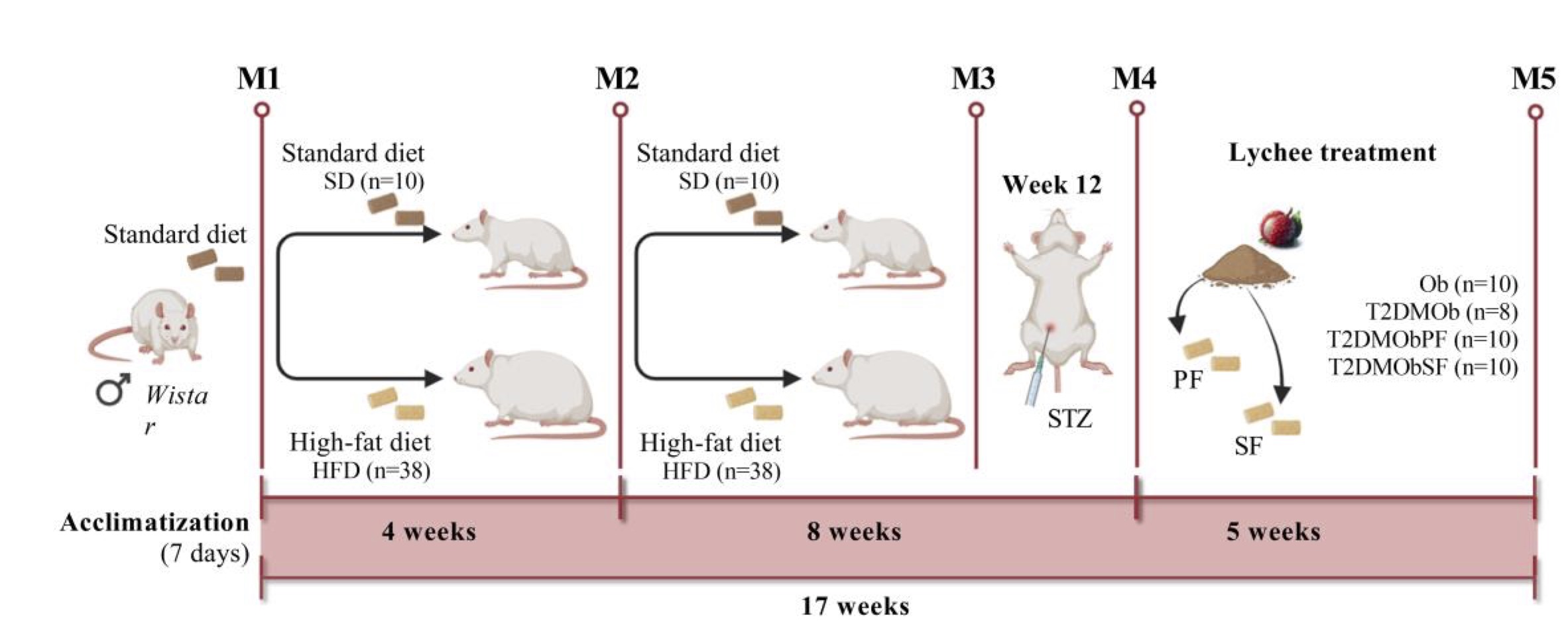
Fig. 1: Schematic representation of the experimental protocol. M1: Obesity induction (4 weeks); M2: Maintenance to Obesity (8 weeks); M3: Type 2 Diabetes Mellitus (T2DM) induction (week 12) with intraperitoneal injection of streptozotocin; M4: Obesity and T2DM maintenance and treatments with litchi peel and seed flours (5 weeks); M5: End of experimental protocol, euthanasia and post mortem analysis. SD: Standard diet; HFD: High-fat diet; Ob: Obese; T2DMOb: Type 2 Diabetes Mellitus obese; T2DMObPF: Type 2 Diabetes Mellitus obese with Lychee peel flour; T2DMObSF: Type 2 Diabetes Mellitus obese with Lychee seed flour; PF: Lychee peel flour; SF: Lychee seed flour; STZ: intraperitoneal injection of streptozotocin.
Initial Moment of Obesity
Obesity was determined from the weekly evolution of the rat’s body weight. This procedure was performed to verify the initial moment and duration of obesity. Previous studies from our research group showed a statistical difference in body weight between these experimental groups (HFD>SD), characterizing the initial moment of obesity [16-18]. This difference in body weight was confirmed when the HFD animals had a significantly higher body mass compared to the SD group. This moment was regarded as the beginning of obesity (week 4). After this moment, SD and HFD rats were maintained in their respective groups for 8 consecutive weeks. In the 12th week, a 95% confidence interval was applied based on the mean body weight of the SD and HFD rats and was adopted as the separation point or midpoint between the lower limit of HFD and the upper limit of SD [16]. Thus, the animals from the HFD group were characterized as obese. The animals in the SD group only remained in the study until the 12th week, with the aim of making them a parameter for the identification of obesity (Fig. 1).
Induction of Type 2 Diabetes Mellitus and Randomization of Treated Groups
In the 12th week, the HFD group was randomized into two groups: obese (Ob, n = 10) and type 2 diabetes mellitus obese (T2DMOb, n = 28). The T2DMOb group were given an intraperitoneal (i.p.) injection of streptozotocin (STZ) (30 mg/kg/i.p.) diluted in 0.1 M citrate buffer (pH 4.5) for induction of T2DM. After 72 hours of STZ administration, animals with fasting blood glucose levels greater than 200 mg/dL were considered T2DM [19]; fasting blood glucose was measured with an Accu-Chek Go Kit portable glucometer (Roche Diagnostic Brazil Ltd., São Paulo, Brazil). Animals that did not respond positively to the dose of STZ were subjected to a second application. The Ob group received the same dosage of a sodium citrate solution.
At the end of week 12, the T2DMOb group was randomized into three new groups: 1) T2DMOb, fed an HFD; 2) T2DMObPF, fed an HFD with added lychee peel flour; and 3) T2DMObSF, fed an HFD with added lychee seed flour. At that moment, the treatment period with lychee peel and seed flours began (5 weeks), lasting until the end of the protocol (Fig. 1). In addition, the Ob group was maintained as the control of T2DMOb.
Lychees and Flour-Making
Lychee fruits (100 kg) were purchased on a rural property located in the region of Marechal Floriano, Espírito Santo, Brazil. Then, they were washed, sanitized with sodium hypochlorite for 15 minutes, and separated into peel, pulp, and seed. The peel and seed were dried in an oven (MA035/5, Drying Oven with Circulation, Marcone, Piracicaba). The temperature used was 45°C until they were a constant weight; it took four days for the lychee peel to dry and eight days for the lychee seed [9]. After drying, they were ground separately in a knife mill (MA340, Macro Knife Mill, Type Willey, Marcone, Piracicaba), and the lychee peel (PF) and seed (SF) flours were obtained, which were stored under refrigeration and protected from light until their use in the formulation of animal feed [9]. All the procedures mentioned above were performed at the Laboratory of Mechanization and Precision Agriculture in the experimental area of the Federal University of Espírito Santo campus in the city of Alegre, Espírito Santo, Brazil.
The HFD was used as a basis for formulating diets with the addition of lychee flour (HFD plus PF and HFD plus SF). Experimental diets with lychee flour were added with 10% PF or SF, corresponding to the amount of flour used in other studies [9, 20]. To balance the possible effect of the amount of fiber present in the PF and SF flours, HFD was added with insoluble fiber during the treatment period with the experimental diets [9].
Total Phenols and Flavonoids in Lychee Peel and Seed Flours
To determine the total phenolic content (TPC), the methodology of Neves et al [21]. and Singleton & Jr [22] was used. 125 μl of the 10% Folin-Ciocalteau reagent aqueous solution and 25 µl of the sample stock solution (1 mg/mL diluted 1:3 in distilled water) were added to a microplate. The mixture was allowed to stand for 5 minutes. After this period, 100 µl of 4% sodium carbonate aqueous solution was added. The reading was performed at a wavelength of 750 nm, in a microplate reader (BioRad, Washington, USA), after approximately 2 hours of incubation in the dark. Gallic acid was used as a standard for the calibration curve and for the expression of the result, which was given in milligrams of gallic acid equivalent per gram of the sample (mgGAE/g).
The total flavonoid content (TFC) was determined by the methodology of Marques et al [23]., with adaptations. In a microplate, 99 μl of distilled water, 20 μl of extract (with final concentration of 100 μg/ml), 6μl of glacial acetic acid, 100 μl of 20% pyridine, and 25 μl of 6.5% aluminum chloride diluted in methanol were added. After 30 minutes, a spectrophotometric reading was performed in a microplate reader (BioRad, Washington, USA) at 415 nm. The reaction blank was considered using a solution with all reagents, excluding the sample and aluminum chloride. The negative control consisted of all reagents except aluminum chloride. Quercetin (Sigma, Missouri, USA) was used as the standard for the calibration curve, and the results were expressed in milligram quercetin equivalent (QE) per gram of sample (mgQE/g).
Table 1 shows the total average of phenolic and flavonoid compounds present in the lychee seed and peel, respectively.
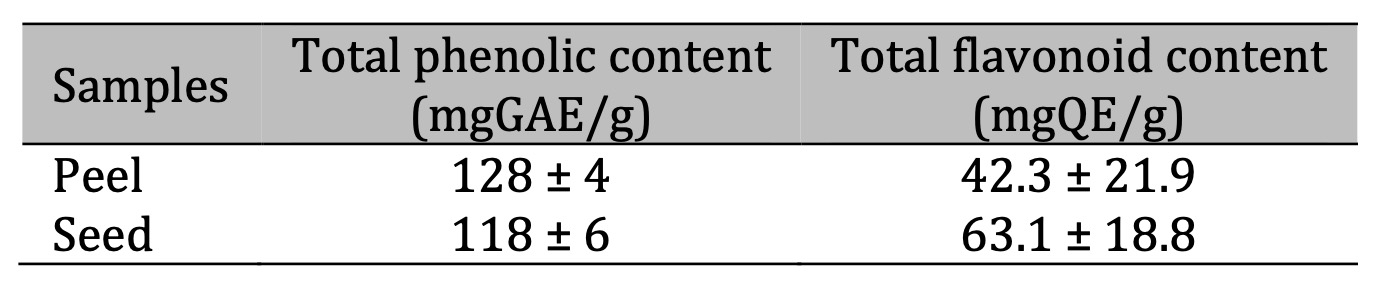
Table 1: Total phenolic and flavonoid in lychee peel and seed flours. GAE: gallic acid equivalent; QE: quercetin equivalent
Euthanasia
At the end of the experimental protocol (17 weeks), the animals received a solution containing ketamine hydrochloride (70 mg/kg/i.p.) and xylazine hydrochloride (10 mg/kg/i.p.) In cases where the animals still exhibited signs of nociceptive reflex after anesthetic induction, an anesthetic overdose (lethal dose) was administered, which consisted of three times the doses of ketamine hydrochloride and xylazine hydrochloride used during the animal’s anesthetic induction [24]. After euthanasia, the animals underwent median thoracotomy; blood samples were collected in Falcon tubes and centrifuged at 3, 000 rpm for 10 minutes (Eppendorf1Centrifuge 5804-R, Hamburg, Germany), and the serum was stored in a freezer at -80˚C (Thermo Fisher Scientific LLC, Asheville, NC, USA). Adipose and cardiac tissue were dissected and collected. Tibia were dissected and measured for later normalization of the heart weight.
Adiposity Characteristics
Body fat was measured by adding the fat deposits in the epididymal, retroperitoneal, and visceral (mesenteric) regions. The adiposity index (AI) was calculated by dividing the total body fat by the final body weight and multiplying by 100 [25].
Biochemical Analyzes
Blood samples were collected in Falcon tubes, centrifuged at 3000 rpm for 10 minutes, and stored in a freezer at -80˚C. Serum concentrations of cholesterol, triglycerides, high-density lipoprotein (HDL), low-density lipoprotein (LDL), and glucose were determined using specific kits (Bioclin ®, Brazil and Synermed do Brasil Ltda, Brazil) and analyzed by the biochemical analyzer automated BS-200 (Mindray Brazil). Serum concentrations of insulin (Linco Research Inc, St. Louis, MO, USA) were quantified by the enzyme-linked immunosorbent assay method (ELISA). Insulin resistance was measured by serum concentrations of glucose and fasting insulin, using the HOMA-IR index (homeostatic model assessment) as proposed by Matthews et al. [26].
Cardiac Remodeling
Cardiac remodeling at the macroscopic level, which identifies the presence or absence of cardiac hypertrophy, was determined by analyzing the following parameters: heart and heart normalized by tibia length [27].
Biomarkers of Oxidants and Antioxidants
The samples were stored in a freezer at -80˚C for later analysis. Then, the samples were sent to São Paulo State University (UNESP), São Paulo, Brazil. Analysis of biomarkers of oxidants and antioxidants was subsequently performed at the RedOx e Inflammation Laboratory at UNESP. Dosages of malondialdehyde (MDA) and carbonylated protein, biomarkers of oxidative stress, were evaluated in serum. The concentrations of malondialdehyde (MDA) for membrane lipid peroxidation evaluation were performed using the thiobarbituric acid reactive substances (TBARS) method [28, 29]. The carbonylated proteins were measured using a nonspecific method that consists of using the 2, 4-dinitrophenylhydrazine (DNPH) derivative and spectrophotometric detection of any proteins modified by the carbonylation process [30]. Concentrations of carbonylated proteins was expressed as nmol DNPH/mg of protein, being the total protein concentrations measured by the Bradford method.
The antioxidant enzyme activities of superoxide dismutase (SOD) and catalase (CAT) were evaluated in serum. SOD activities were determined based on the inhibition of a superoxide radical reaction with pyrogallol [31]. Changes in absorbance of the reaction solution at 420 nm were determined after 1 min. The values are expressed as units per milligram of protein per minute. CAT activities were evaluated by following the decrease in the levels of hydrogen peroxide. The absorbance values were measured at 240 nm and expressed as µmol/mL/min [32].
Cardiomyocyte Contractility
Cellular contractile was measured using the cardiomyocyte length change technique with an edge detection system coupled to an inverted microscope (IonOptix, Milton, MA, USA) with a 40x objective lens (Nikon Eclipse - TS100, USA). The cardiomyocytes were placed in an experimental camera with a glass base, bathed in Tyrode solution, and viewed on a monitor using a camera (Myocam, IonOptix, Milton, MA, USA, 240 Hz) coupled to the microscope with an image detection program (Ionwizard, Ionoptix, Westwood, Massachusetts, USA).
Cardiomyocytes were stimulated at a frequency of 1 Hz lasting 5 milliseconds and a voltage of 20V, using a pair of steel electrodes and an electrical field stimulator (Myopacer, IonOptix, Milton, MA, USA). Contractile measurements were performed on isolated cardiomyocytes that presented the following conditions: well-defined sarcomere borders and striations, relaxed at rest, and not presenting voluntary contractions. Fractional shortening (expressed as a percentage of resting cell length), maximal rates of shortening and relaxation, and times to 50% shortening and relaxation were measured. The total numbers of cells analyzed are described in the legend of each figure.
Intracellular Ca2+ Measurements
Myocytes were loaded with 1.0 μM Fura2-acetoxymethyl (AM) ester (Molecular Probes, Eugene, OR, USA) for 10 minutes at room temperature, washed with Tyrode solution, and allowed to rest for an additional 10 min to allow the de-esterification of the dye. Subsequently, the cardiomyocytes were stimulated at 1 Hz (Myopacer 100, Ionoptix Inc.), and fluorescence images were obtained using excitation of 340 to 380 nm wavelengths using a Hyper Switch system (IonOptix, Milton, MA). Background-subtracted fluorescence emission was obtained, and the Fura 2AM ratios were used as an index of intracellular [Ca2+]i transient, which detected at approximately 510 nm. The Ca2+ amplitude was reported as F/F0, where F is the maximal fluorescence intensity average measured at the peak of [Ca2+]i transients, and F0 is the baseline fluorescence intensity measured at the diastolic phase of [Ca2+]i transients. Systolic and diastolic Ca2+ times 50% peak Ca2+ and 50% Ca2+ decay were also analyzed. In addition, the ratios of systolic Ca2+ by shortening fraction and by maximum rate of shortening were performed to evaluate the responsiveness of myofilaments to Ca2+.
Statistical Analysis
Data are displayed as mean ± standard deviation, and a Shapiro–Wilk normality test was used for normality evaluation. For comparison between two groups, Student’s t test and Mann-Whitney test were carried out for parametric and nonparametric data, respectively. BW evolution was evaluated using two-way repeated measures ANOVA, complemented with a post-hoc Bonferroni test. Comparisons between the three groups were performed by one-way ANOVA, complemented by Tuckey or Kruskal-Wallis post-hoc tests. The association between fractional shortening and pro-oxidant and antioxidant markers was found using Pearson’s correlation coefficient. The significance level considered was 5%. The statistical analyses and graphics were conducted using GraphPad Prism 9.0 software (GraphPad, San Diego, CA, USA).
Results
Fig. 2 presents the weekly body weight progression of the groups throughout the experimental period (17 weeks). At the beginning of the experimental protocol, the animals had similar body weights, but the initial moment of obesity was observed in week 4, since the HFD animals showed a higher body weight compared to the SD group, and this was maintained until the 12th week. In addition, the presence of T2DM promoted a reduction of body weight gain when compared to nondiabetic animals (T2DMOb < Ob). Furthermore, no changes were observed in the body weight of T2DMOb animals during the period of treatment with lychee peel and seed flours (Fig. 2).
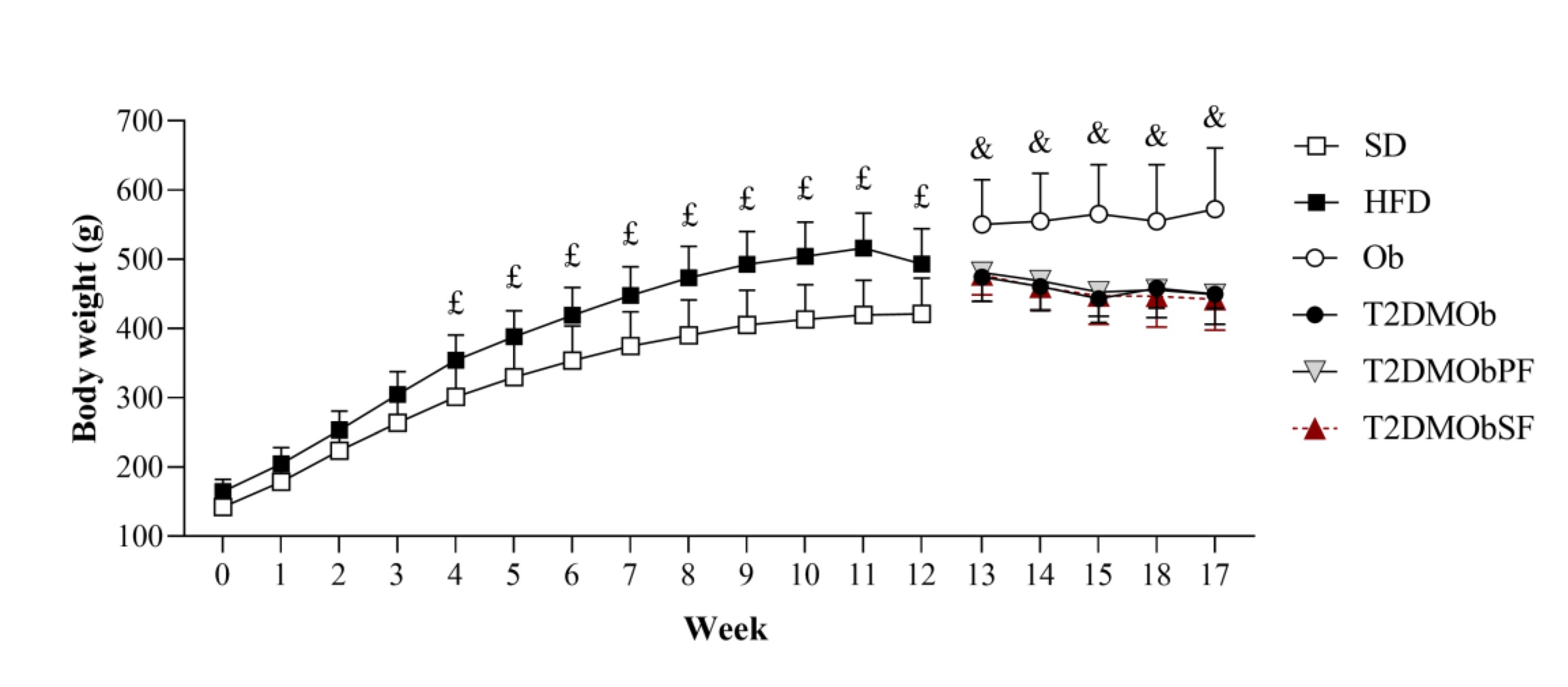
Fig. 2: Evolution of body weight. Weeks 0 to 12: standard diet (SD, n=10); high-fat diet (HFD, n=38). Weeks 13 to 17: obese (Ob, n=10); type 2 Diabetes Mellitus obese (T2DMOb, n=8); type 2 Diabetes Mellitus obese with litchi with peel flour (T2DMObPF, n=10); type 2 Diabetes Mellitus obese with lychee seed flour (T2DMObSF, n=10). Data are presented as mean ± standard deviation. Two-way repeated measures ANOVA complemented by Bonferroni's post-hoc test, p<0.05- £SD vs. HFD; &Ob vs. T2DMOb.
Glycemic levels in pre- and post-induction of T2DM with intraperitoneal injection of STZ are shown in Fig. 3. There was no statistical difference in glycemic levels in the Ob group before and after sodium citrate solution application, as well as between the Ob and T2DMOb groups in the pre-drug moment. However, T2DMOb animals had a higher glycemic level between the pre- and post-application of the drug; the same result was observed between the T2DMOb and Ob post-STZ/citrate groups, characterizing the onset of diabetes in the T2DMOb group.
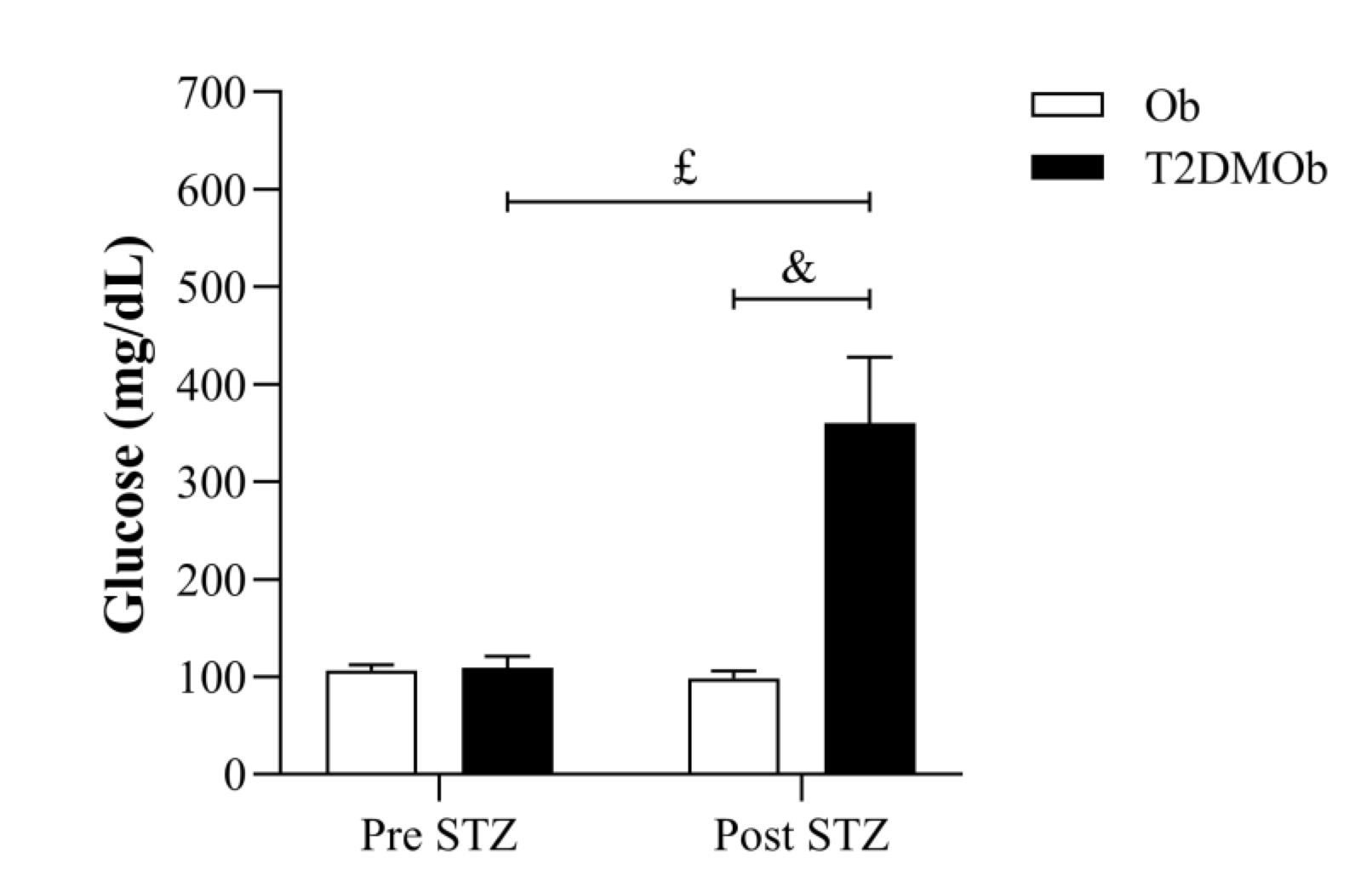
Fig. 3: Glycemic level in pre- and post-induction of type 2 diabetes mellitus (T2DM) with intraperitoneal injection of streptozotocin (week 12). Ob: obese group was presented as control of T2DMOb (n=10); T2DMOb: Type 2 Diabetes Mellitus obese (n=28). STZ: streptozotocin. Data are presented as mean ± standard deviation. Student's t-test, p<0.05- &Ob vs. T2DMOb. Two-way repeated measures ANOVA complemented by Bonferroni's post-hoc test, p<0.05- £Pre STZ vs. Post STZ.
The nutritional profile and general characteristics of the groups are shown in Table 2. The T2DMOb animals had a higher food consumption and caloric intake compared to the Ob group; however, they had lower feed efficiency, which justifies the lower body weight gain compared to the Ob group (Fig. 2). The lower body weight of the T2DMOb group was accompanied by lower adiposity, presenting a reduction of epididymal, visceral, and retroperitoneal fat pads, total body fat, and adiposity index when compared to the Ob group. No statistical difference was found in heart weight and heart/tibia length with the presence of T2DM (Table 2).
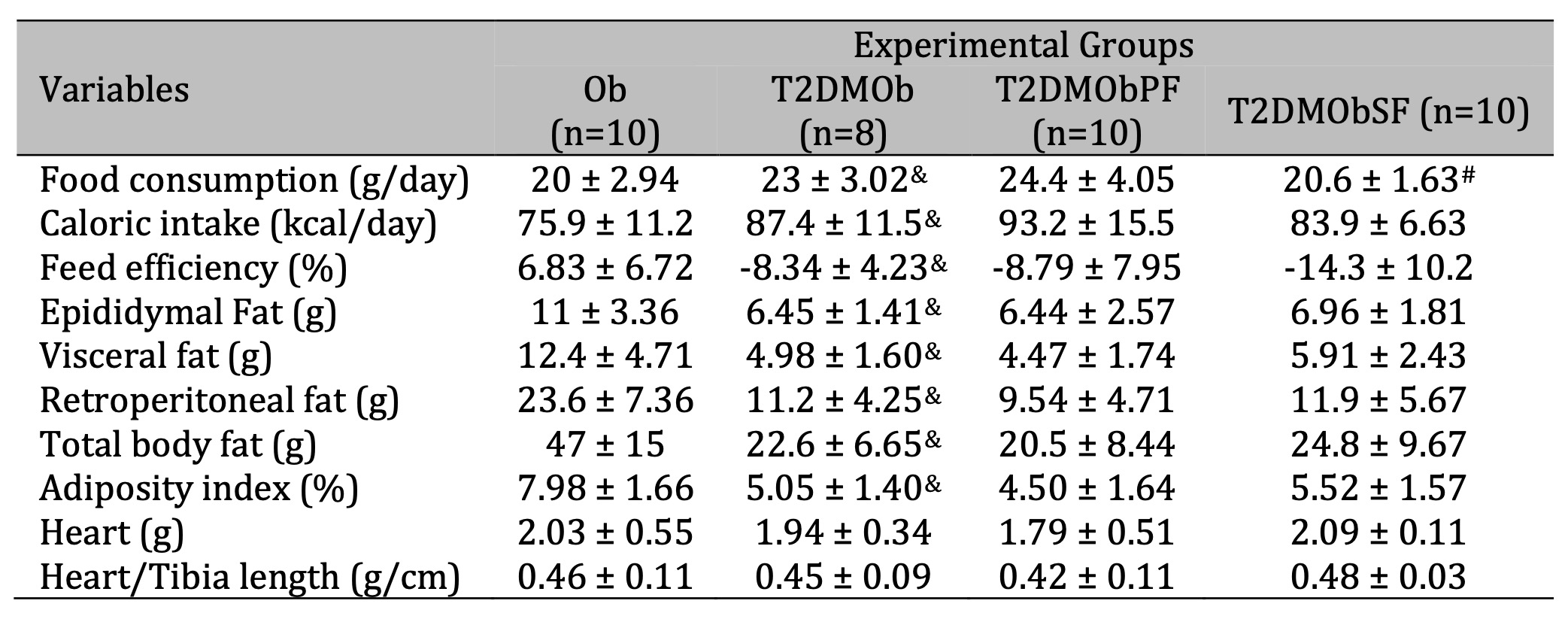
Table 2: Nutritional profile and general characteristics. Ob: obese group was presented as control of T2DMOb; T2DMOb: Type 2 Diabetes Mellitus obese; T2DMObPF: Type 2 Diabetes Mellitus obese with lychee peel flour; T2DMObSF: Type 2 Diabetes Mellitus obese with lychee seed flour. IBW: initial body weight; FBW: final body weight. Data are presented as mean ± SD. Student's t-test, p<0.05 - & Ob vs. T2DMOb; For comparison of T2DMOb, T2DMObPF and T2DMObSF: One-way ANOVA for independent samples complemented by Tuckey post-hoc test, p<0.05 - # T2DMObSF vs. TT2DMObPF
Considering the treatments of lychee peel and seed flours, the results showed that T2DMObSF had lower food consumption in relation to group T2DMObPF (-15.6%). There was no effect of lychee peel and seed flour treatments on nutritional, adiposity, and cardiac morphologic parameters on T2DMOb (Table 2).
As presented in Fig. 4, the presence of T2DM resulted in elevated serum glucose and HDL levels but a reduction in insulin levels (Figures 4A, B, and F). However, HOMA-IR, cholesterol, triglycerides, and LDL parameters were similar between the groups (Figures 4C, D, E, and G). Additionally, treatments with lychee peel and seed flour did not alter the glycemic, insulinemic, and lipid profiles on T2DMOb.
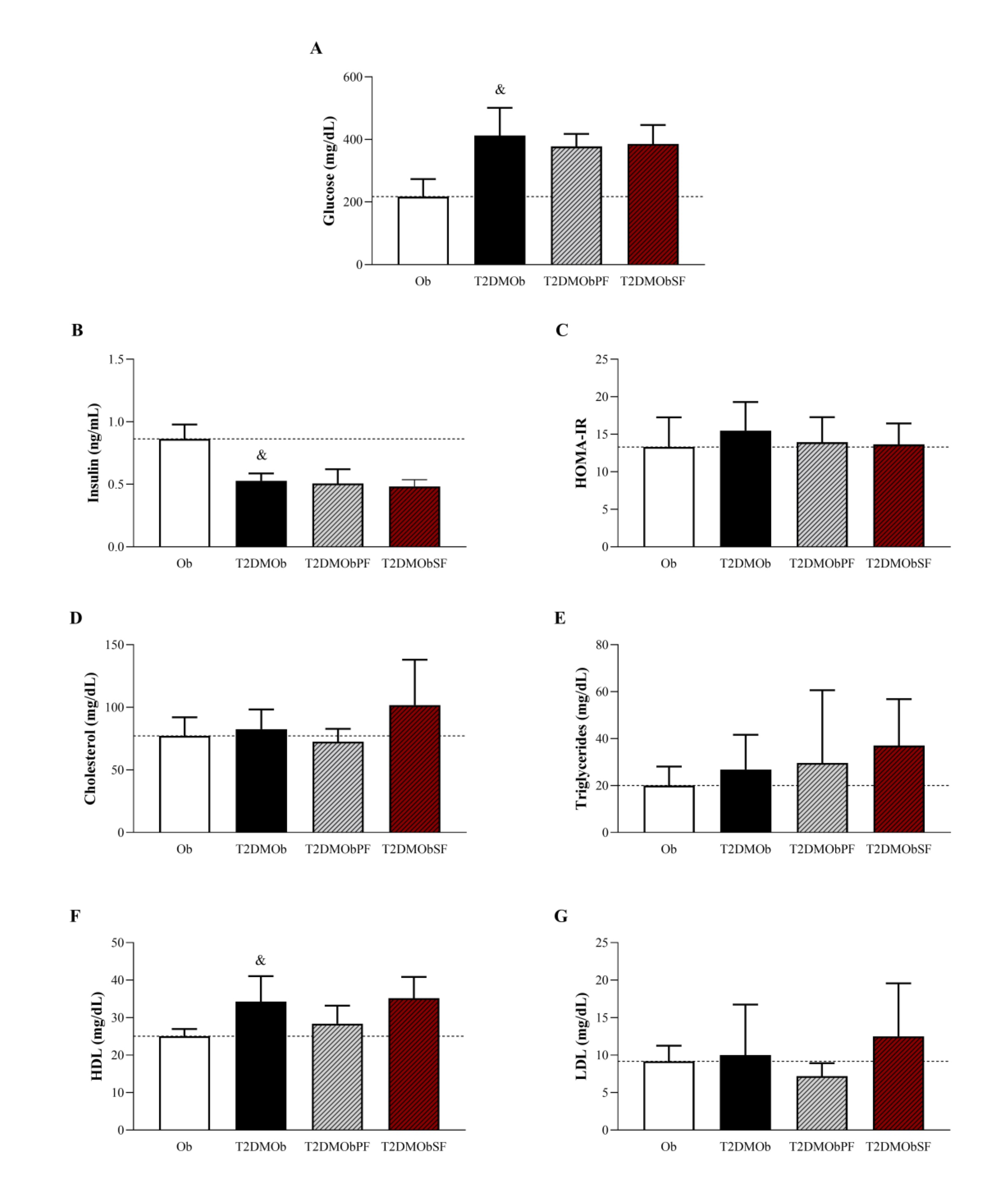
Fig. 4: Glycemic, insulinemic and lipid profiles of Type 2 Diabetes Mellitus obese (T2DMOb) treated with lychee peel and seed flour. Ob: obese group was presented as control of T2DMOb (n=9); T2DMOb: Type 2 Diabetes Mellitus obese (n=8), T2DMObPF: Type 2 Diabetes Mellitus obese with lychee peel flour (n=7) and T2DMObSF: Type 2 Diabetes Mellitus obese with lychee seed flour (n=6); HOMA-IR: homeostatic model assessment index; HDL: high-density lipoprotein; LDL: low-density lipoprotein. Data are presented as mean ± standard deviation. Student's t-test, p<0.05- &Ob vs. T2DMOb. For comparison of T2DMOb, T2DMObPF and T2DMObSF: One-way ANOVA for independent samples complemented by Tuckey post-hoc test.
Fig. 5 shows the results related to oxidant and antioxidant biomarkers. The T2DMOb group exhibited higher serum levels of protein carbonylation and SOD compared to the Ob group (Figures 5B and C). However, no changes in MDA and catalase levels were observed in T2DMOb. Treatments with lychee peel and seed flours were not able to reverse oxidative damage or improve the antioxidant capacity in T2DMOb on T2DMOb, since the concentrations of MDA, protein carbonylation, SOD, and catalase were similar among groups.
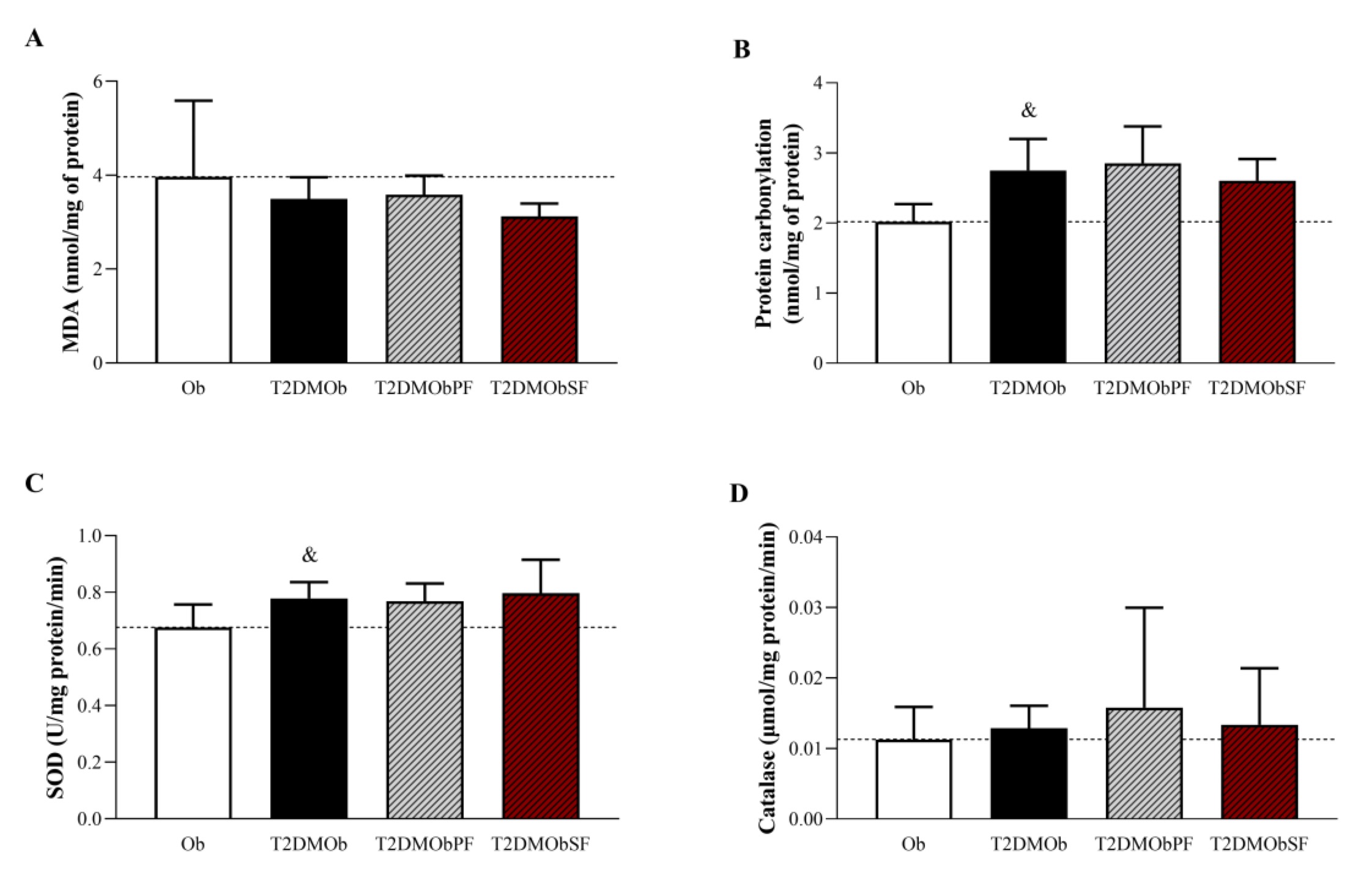
Fig. 5: Oxidative biomarkers of Type 2 Diabetes Mellitus obese (T2DMOb) treated with lychee peel and seed flours. Ob: obese group was presented as control of T2DMOb (n=8); T2DMOb: Type 2 Diabetes Mellitus obese (n=8), T2DMObPF: Type 2 Diabetes Mellitus obese with lychee peel flour (n=7) and T2DMObSF: Type 2 Diabetes Mellitus obese with lychee seed flour (n=7); MDA: Malondialdehyde; SOD: superoxide dismutase; CAT: catalase. Data are presented as mean ± standard deviation. Student's t-test, p<0.05 - &Ob vs. T2DMOb. For comparison of T2DMOb, T2DMObPF and T2DMObSF: One-way ANOVA for independent samples complemented by Tuckey post-hoc test.
T2DM promoted an increase in fractional shortening without alterations in the maximum rate of shortening and time to 50% shortening (Figures 6A, B, and D). Lychee treatments did not accentuate the elevation of fractional shortening in T2DMOb (Fig. 6A). In addition, the treatment with lychee peel and seed flour promoted an elevation of time to 50% shortening when compared to the T2DMOb group (Fig. 6D) without alterations in the maximum rate of shortening (Fig. 6B). Regarding relaxation, the maximum rate of relaxation was higher in the T2DMOb group compared to the Ob group, but the lychee treatments did not promote positive alterations in this parameter (Fig. 6C). However, the T2DMObSF group had a lower maximum rate of relaxation when compared to the T2DMObPF group (Fig. 6C). One important result is related to time to 50% relaxation, which was also longer in the presence of T2DM (T2DMOb > Ob); the treatments with lychee peel flour showed a reduction in the time to 50% relaxation (Fig. 6E). Additionally, when comparing the lychee-treated groups for the same parameter, the T2DMObSF group took a longer time to 50% relaxation than the T2DMObPF group (Fig. 6E).
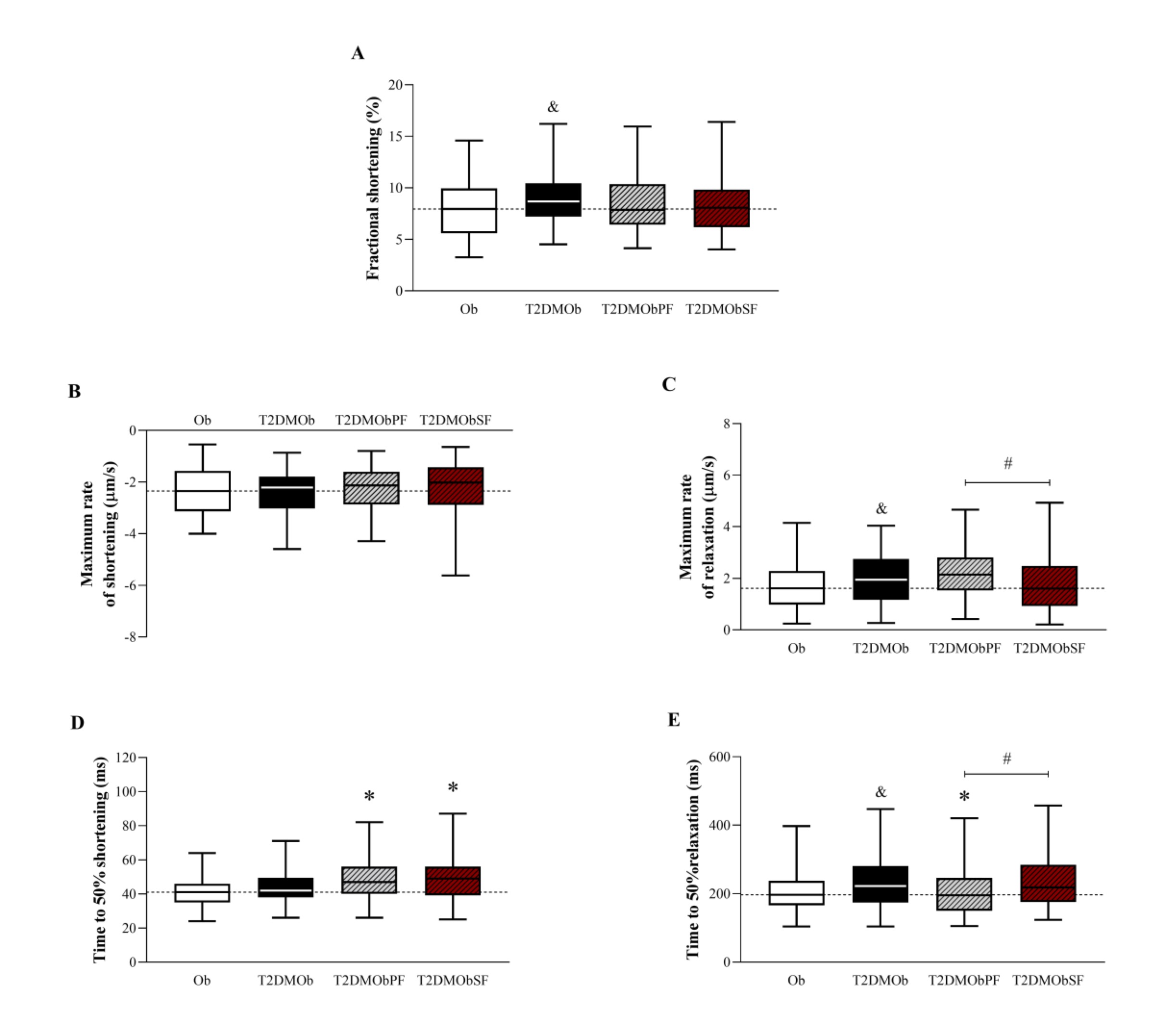
Fig. 6: Functional contractility parameters of isolated cardiomyocytes stimulated at 1 Hz from Type 2 Diabetes Mellitus obese (T2DMOb) treated with lychee peel and seed flours. Ob: obese group was presented as control of T2DMOb (n=6, cells = 100); T2DMOb: Type 2 Diabetes Mellitus obese (n= 5, cells = 101); T2DMObPF: Type 2 Diabetes Mellitus obese with lychee peel flour (n= 5, cells = 99); T2DMObSF: Type 2 Diabetes Mellitus obese with lychee seed flour (n= 7, cells = 92). Data are presented as median ± interquartile range (Figures A, B, D, E) or expressed as mean ± standard deviation (Figure C). Mann-Whitney test, p<0.05 - & Ob vs. T2DMOb; For comparison of T2DMOb, T2DMObPF and T2DMObSF: Kruskal-Walli’s test complemented by Dunn’s post-hoc test. p<0.05 - * vs. T2DMOb; # T2DMObSF vs. T2DMObPF.
Considering the contractile parameters, the results showed that systolic Ca2+ and time to 50% decay Ca2+ were elevated in the T2DMOb in relation to Ob animals, but these parameters were not improved by treatments with lychee peel and seed flour (Fig. 7B and E). In addition, the T2DMOb rats presented an increase in diastolic Ca2+ when compared to the Ob group (Fig. 7C). It is important to note that the treatment with lychee seed flour reduced this parameter, despite not improving the time to 50% decay Ca2+ and, consequently, relaxation (Fig. 7C and E). Time to 50% peak Ca2+ was similar among the groups. The treatments with lychee peel and seed flour did not change the sensitivity of myofilaments to Ca2+ in T2DMOb animals, since the systolic Ca2+/fractional shortening and systolic Ca2+/maximum rate of shortening ratios were similar among groups.
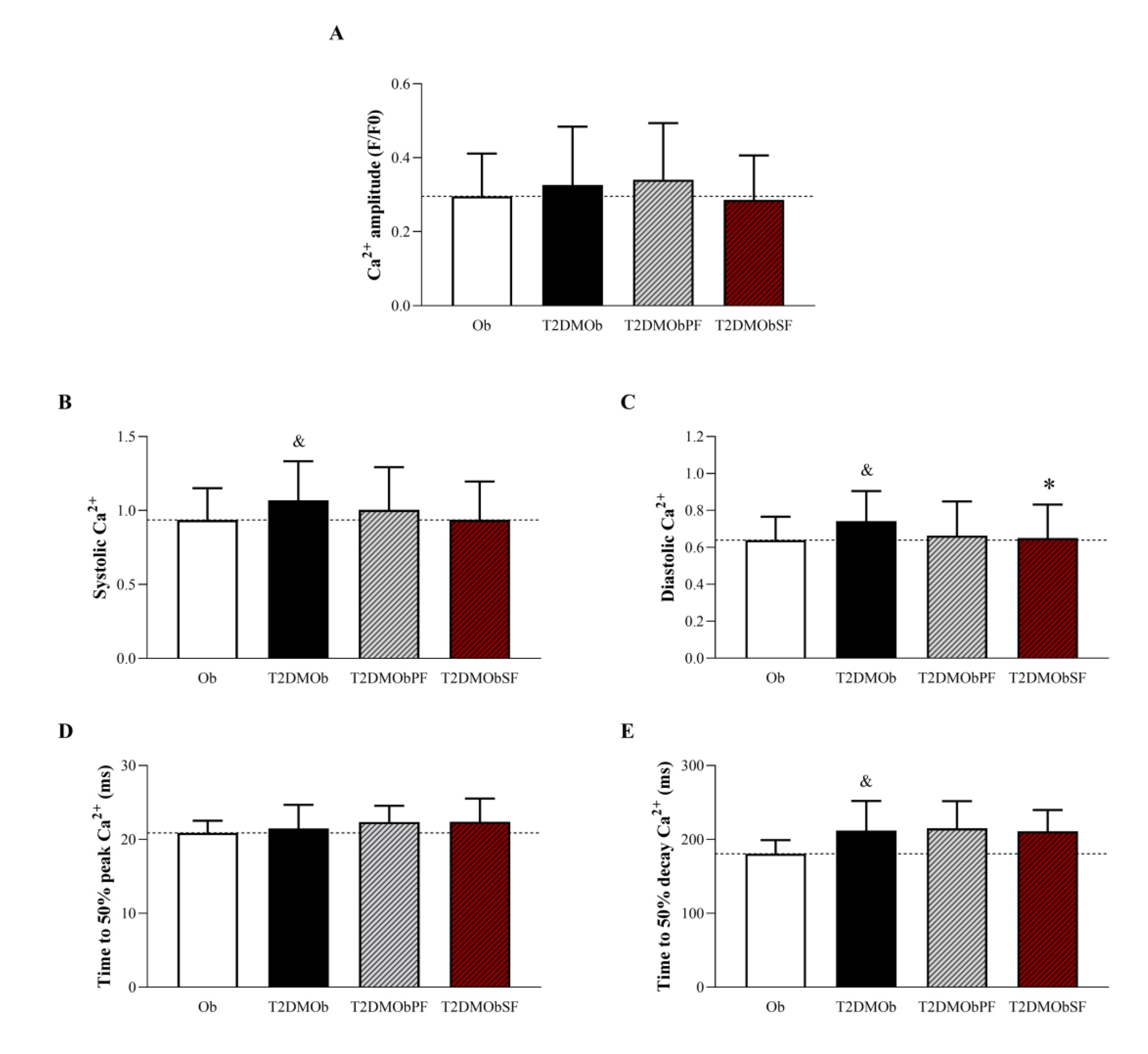
Fig. 7: Intracellular Ca2+ handling of isolated cardiomyocytes stimulated at 1 Hz from Type 2 Diabetes Mellitus obese (T2DMOb) treated with lychee peel and seed flours. Ob: obese group was presented as control of T2DMOb (n=4, cells = 50); T2DMOb: Type 2 Diabetes Mellitus obese (n= 4, cells = 40); T2DMObPF: Type 2 Diabetes Mellitus obese with lychee peel flour (n=5, cells = 39); T2DMObSF: Type 2 Diabetes Mellitus obese with lychee seed flour (n= 4, cells = 45). Data are presented as the mean ± standard deviation. Student's t-test, p<0.05 - &Ob vs. T2DMOb; For comparison of T2DMOb, T2DMObPF and T2DMObSF: One-way ANOVA for independent samples complemented by Tuckey post-hoc test, p<0.05 - * vs. T2DMOb.
Correlations between myocardial contractility (fractional shortening) and pro-oxidant and antioxidant markers are summarized in Fig. 8. A weak correlation was observed between fractional shortening and SOD (r = -0.2135) (Fig. 8C), but it was not significant. Other markers indicate a very weak correlation (Figures 8A, B, and D).
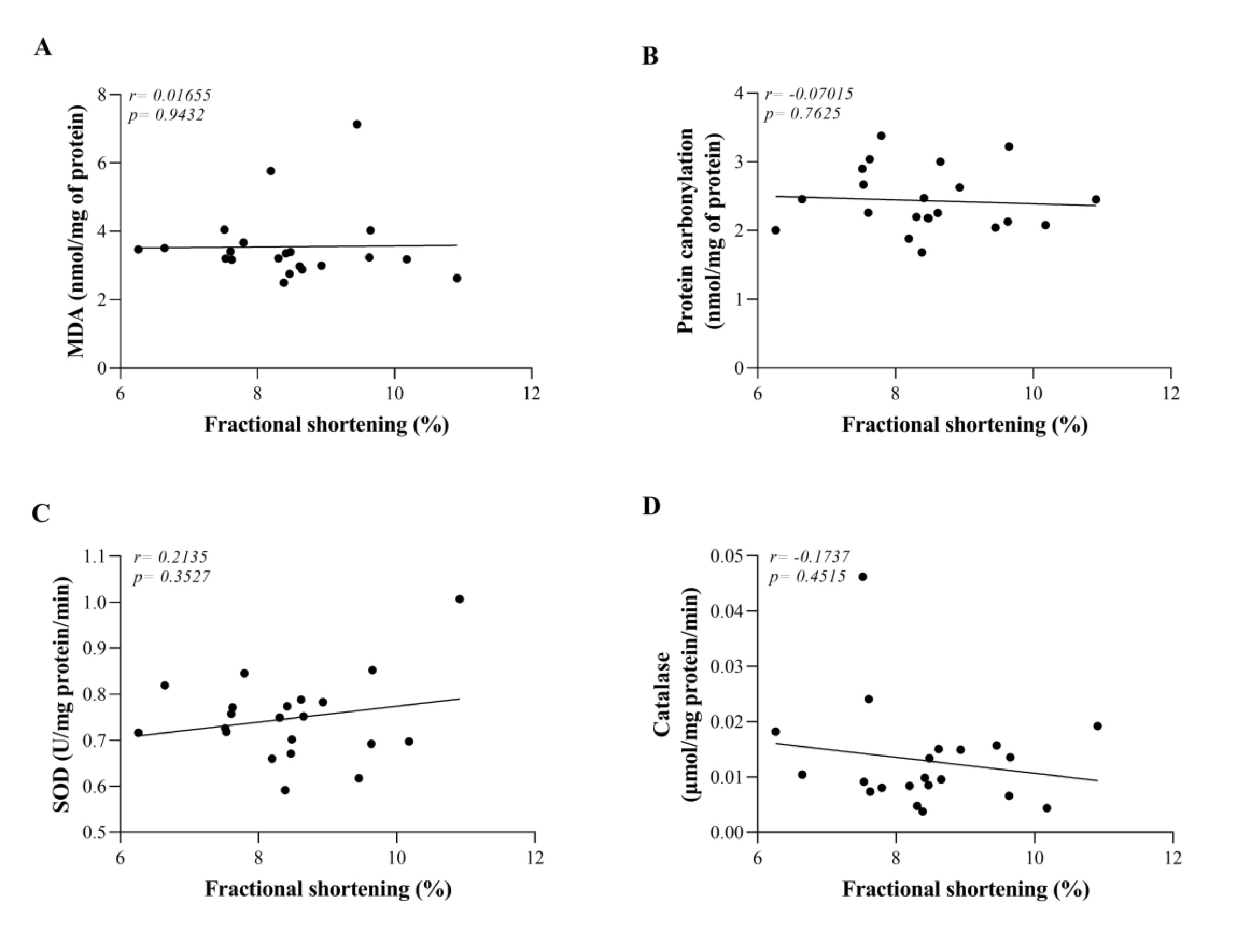
Fig. 8: Correlations between myocardial contractility (fractional shortening) and pro-oxidant and antioxidant markers. Ob: obese group was presented as control of T2DMOb (n=6); T2DMOb: Type 2 Diabetes Mellitus obese (n= 5); T2DMObPF: Type 2 Diabetes Mellitus obese litchi peel flour (n= 5); T2DMObSF: Type 2 Diabetes Mellitus obese litchi seed flour (n= 5). MDA: Malondialdehyde; SOD: superoxide dismutase; CAT: catalase. Data are presented as the mean. Pearson's correlation coefficient.
Discussion
The current study aimed to investigate the effect of lychee peel and seed flour consumption on obese T2DM rats and the effects on anti/pro-oxidant systems and cardiomyocyte contractile function. The main finding of the study was that the treatment with lychee peel flour favored a reduction in the time to 50% relaxation, which was impaired in the T2DM condition. In addition, lychee peel and seed flours do not reverse oxidative damage in T2DMOb rats.
Regarding the development of obesity, we showed that in the 4th week of obesity induction, the HFD animals showed a significant difference in body weight in relation to group SD; this difference remained throughout the period of maintenance of obesity. Other studies have demonstrated that an HFD is efficient in inducing obesity, leading to a difference in body weight between obese and control groups [16, 33].
After obesity was established, the animals were induced to T2DM by the application of STZ (30 mg/kg of weight), characterizing associated diabetes to obesity T2DMOb). Studies have shown that a low dose of STZ (30 mg/kg) applied once or twice is capable of inducing T2DM in rats submitted to an HFD [11, 34]. The STZ induction model has the advantages of low cost, easy application, and consequent selective damage to pancreatic β cells, as it keeps the others intact, mimicking the metabolic characteristics and natural history of T2DM developed in humans [35, 36].
In T2DM, the body does not adequately use the insulin produced by the pancreas; it is not even able to produce enough insulin to control the blood glucose level [2]. The classic diabetic glycemic profile was found in the current study, in which the T2DMOb group presented high glycemic levels and low insulin levels. However, no differences were noted in the blood glucose levels of the T2DMObPF and T2DMObSF groups when compared to the T2DMOb group; although the treated groups showed a slight decrease in glycemic levels, it was not enough to demonstrate significance. Studies with lychee seed extracts administered to T2DM animals observed a decrease in blood glucose, insulin, and the insulin resistance index [11, 37]. Furthermore, Queiroz et al [20]. observed a reduction in blood glucose levels in animals that received lychee peel flour; their results were attributed to the fiber present in this flour.
An explanation for our findings can be related to the amount of polyphenols in a high-fat diet. Polyphenols are secondary metabolites of plants that are responsible for several functions, being antiviral, antimicrobial, and antioxidant and also modulating physiological processes in plants [38]. It is important to note that in foods, they are responsible for nutritional and organoleptic characteristics [38], being located in the vacuoles of plant cells protected by the cell wall [39]. The cell wall is ruptured during chewing, leaving these substances available for absorption; however, not all cells are ruptured [39]. In the current study, considering the food consumption of animals and the concentration of polyphenols in lychee peel and seed flour, the amount of polyphenols in the seed flour was reduced, representing only 3.5% of the amount offered by Man and collaborators [11] in seed extract. Furthermore, in a study carried out by Kilari and Putta [12], the administration time of lychee peel extract was 12 weeks, while in the present study, the animals received the diet with the flours for a period of 5 weeks. Tang and collaborators [37] also point out that some results found by the seed extract as an index of insulin resistance are dose dependent; therefore, the higher the dose, the better the result. In the current study, the dose of flour was that suggested by Queiroz and collaborators [20]; however, the flour administration time was longer.
As presented, the T2DM model used in the current study was efficient in inducing some metabolic changes present in human diabetes [40], such as glucose intolerance, hypoinsulinemia, and change in adiposity. However, an unexpected increase in HDL was also observed, which can be attributed to structural and functional modifications of the HDL particle in a dysmetabolic environment. While HDL is traditionally considered protective against cardiovascular disease, in pathological conditions such as T2DM, HDL may undergo alterations that transform its anti-atherosclerotic and antioxidant properties into pro-inflammatory and pro-atherogenic characteristics [41]. In T2DM, HDL can accumulate components such as oxidized fatty acids, resulting in a dysfunctional particle with reduced capacity to promote cholesterol efflux from cells, thus contributing to increased cardiovascular risk. These alterations in HDL may therefore not only reflect changes in lipid and glucose metabolism but also serve as an indicator of the exacerbated inflammatory and oxidative environment characteristic of type 2 diabetes [42, 43]. Previous studies demonstrated hypolipidemic and antiobesity effects with lychee treatment [44, 45]. Su and collaborators [44], when investigating the effect of phenolic-rich extract of lychee pulp in mice fed with HFD, observed an improvement in the lipid profile and reduction in adiposity due to lower fatty acid synthesis and promoting fatty acid β-oxidation. Mhatre, Bhagit, and Yadav [45] reported an anti-obesity effect from the in vitro inhibition of pancreatic lipase due to the action of a protein present in the seeds of the Litchi chinensis fruit [45]. However, there was also no evidence of changes in body weight or serum levels of cholesterol and triglycerides in the STZ/HFD induced T2DM model treated with lychee seed extract, even at doses higher than those used in the diets in the present study, as previously mentioned [11]. This suggests that even at higher doses, the bioactive compound may not affect lipid metabolism and body weight in T2DM models.
Increased oxidative stress in T2DM can be attributed to mitochondrial dysfunction and chronic hyperglycemia. Excess glucose induces an increase in ROS due to several mechanisms, such as glucose auto-oxidation and the development of advanced glycation end-products [46]. ROS can react with amino acids susceptible to oxidation (especially arginine, proline, lysine, and threonine residues), resulting in the formation of carbonyl groups [47]. Protein carbonylation is a nonenzymatic oxidative post-translational modification and thus seriously affects protein functions [48]. Due to its early formation and stability, carbonyl protein is considered a good biomarker of oxidative stress [49]. In the current study, serum levels of carbonyl protein were higher in the T2DMOb group when compared to the Ob group. This result may suggest an elevation of oxidative biomarkers in T2DMOb animals, even without changes in serum concentrations of MDA. Also, considering glycemic levels, it can be inferred that hyperglycemia may have contributed to the high levels of protein carbonyl in these animals. The increase in SOD concentration observed in the current study is conflicting, since antioxidant activity is expected to be reduced in the T2DM condition [50]. Promyos and collaborators [51] described that high and constant levels of SOD may be the result of overexpression of the enzyme as a form of defense against oxidative stress. In addition, these increases can be associated with several factors, including the duration and stage of T2DM [51].
Regarding lychee treatment, there were no differences between the T2DMObPF and T2DMObSF treated groups compared to the T2DMOb group. A study carried out with hypercholesterolemic animals shows that treatment with lychee peel flour resulted in a decrease in TBARS levels in the liver, which was possibly attributed to the presence of antioxidants in the flour [20]. Yang and collaborators [52], when evaluating the effect of gavage administration of lychee peel extract in a genetic model of T2DM, observed an improvement in oxidative stress resulting from reduced levels of MDA and increased levels of SOD. The authors concluded that the improvement in the antioxidant parameter was due to the translocation of nuclear factor erythroid2-related factor 2 into the cell nucleus, promoted by the extract, which contributed to the transcription of SOD [52]. Kilari and Putta [12] demonstrated that the administration of lychee peel extract in STZ-induced diabetic rats reduced blood glucose and showed an increase in the antioxidant enzymes SOD and catalase and a decrease in carbonyl protein; the authors attributed the results to the polyphenols present in the peel extract.
Researchers have stated that lychee peel is an important source of bioactive compounds, such as flavonoids; thus, lychee peel extract has demonstrated antitumor effects [53]. Ethanoic extract of lychee seed also has good antioxidant activity [54]. Although several studies highlight the good antioxidant capacity of lychee peel and seed [12, 20, 53, 54], the current study was not able to promote an improvement in improve oxidative stress of animals induced to T2DMOb, visualized by elevation in protein carbonylation. These results can be attributed to low doses of antioxidant compounds in the amount of flour offered to the animals and/or the lower availability of phenolic compounds of these nutrients compared to lychee peel and seed extracts.
Cardiac remodeling has been extensively investigated in models of diet-induced obesity in our laboratory [15, 27, 55-57]. Changes in the heart occur due to aggression or conditions of prolonged overload, resulting in microscopic changes that translate into changes in the size, mass, shape, and functioning of the heart [58]. Nevertheless, as in the current study, previous studies by our research group also did not show cardiac remodeling visualized by hypertrophy [15, 27, 56, 57], even with the use of different high-calorie diets [15, 56]. Diabetes may be involved in a series of abnormal biochemical events that may contribute to cardiac remodeling, such as Ca2+ signaling, oxidative stress, inflammation, and glucose metabolism [59]. A study carried out by Kishi and collaborators [60] evaluated the cumulative effect over 25 years of exposure to glycemic abnormalities on LV remodeling in young adults. The authors observed that glycemic abnormalities negatively affected cardiac remodeling [60], indicating, then, that changes in the structure of the heart only appear after a prolonged period of stable hyperglycemia [61]. It is thus suggested that the protocol time (17 weeks) was not effective in inducing cardiac remodeling, visualized by the lack of change in heart weight in T2DM, even with the presence of hyperglycemia. Therefore, in the absence of cardiac remodeling, it is expected that this parameter was not attenuated with lychee treatments, as seen in the current study.
Regarding the contractile functional parameters of cardiomyocytes and intracellular Ca2+ handling, the T2DMOb group presented a higher concentration of systolic Ca2+ and increased shortening of cardiac myocytes. As mentioned by Bers [62], Ca2+ is essential for the cardiac excitation-contraction coupling process, and incorrect handling of this ion can lead to contractile dysfunction. Although previous studies have observed impairments in Ca2+ handling in diabetic conditions [63, 64], the T2DM model in the current study showed higher systolic Ca2+ concentration. Howarth and collaborators [64] pointed out that the increase in Ca2+ handling is linked to changes in the sensitivity of the myofilament to Ca2+. After 18 months of age, diabetic animals showed unchanged amplitude of myocardial shortening, at the expense of increased transient Ca2+, which suggests a compensatory mechanism in the face of decreased sensitivity to myofilament Ca2+ [64]. In the current study, there was an increase in the fractional shortening in response to the increase in systolic Ca2+, which suggests that the experimental protocol time was not sufficient to promote changes in calcium sensitivity.
On the other hand, considering the diastolic parameters, the T2DMOb showed a higher maximum rate of relaxation; however, this improvement was not accompanied by an increase in Ca2+ reuptake, since there was an increase in diastolic Ca2+ levels and a longer time until 50% of Ca2+ reuptake. This result is in agreement with a cohort study that shows diastolic dysfunction resulting from an increase in Ca2+ in the cytosol, which could explain the impairment in relaxation frequently seen in diabetes [65]. Oxidative stress can promote disturbances in the functioning of Ca2+-related proteins such as SERCA2a [66]. Xu and collaborators [67] showed that ROS can directly inhibit the Ca2+ reuptake function of Serca2a. Thus, the diastolic dysfunction in the current study observed in the T2DMOb group may be due to greater oxidative stress in this condition. The authors have shown that the assessment of protein carbonylation is essential for visualizing oxidative damage in cardiac proteins, being altered in cardiomyocytes from genetically obese rats [68].
There is a scarcity of studies investigating the effect of lychee on myocardial functional parameters in a T2DM model. An in vitro study investigated the effects of phlorizin on contractility and Ca2+ transport in STZ-induced diabetic rat ventricular myocytes [69]; phlorizin is a phenolic compound found in lychee seeds [70]. The authors observed a decrease in the amplitude of the Ca2+ handling and a damage of shortening in myocytes, without altering the Ca2+ myofilament sensitivity. These findings suggest that changes in Ca2+ transport may be partially underlying the reductions in myocyte shortening [69]. A study conducted with Wistar rats evaluated the effect of oral administration of 40 mL/day of green tea for 4 weeks. Green tea is rich in polyphenols, and the main component is epicatechin-3-gallate [71], a flavonoid also present in lychee seeds [70]. Bocchi and colleagues observed an improvement in mechanical properties and Ca2+ dynamics, demonstrating increased Ca2+ release associated with an increased rate of Ca2+ handling decay, without changes in Ca2+ sensitivity or the expression of proteins involved in contraction processes [71].
Quercetin-3-O-rutinoside (rutin) is the predominant flavonoid in lychee cultivar fruits [72] and can be found in both the pulp and peel of lychee [70]. Huang and colleagues [73] evaluated the effect of rutin doses on cardiac function in diabetic ApoE knockout mice over 6 weeks (60 mg/kg body weight). They observed that rutin reduced oxidative damage, hypertrophy, and fibrosis as well as promoting recovered systolic and diastolic functions of diabetic hearts, resulting in overall improvement in myocardial function [73]. Guimaraes and colleagues [74], in their evaluation of the effect of administering 50 mg/kg body weight for seven days in diabetic rats, found that rutin prevented cardiac remodeling and preserved systolic function impaired by STZ-induced diabetes mellitus.
Taken together, the data suggest that the absence of changes in contractile parameters is justifiable, given that there was no impairment in contractility in the T2DM model used in the present study. On the other hand, the bioactive compounds present in lychee peel were effective in promoting faster cardiac relaxation.
In conclusion, treatments using lychee seed and peel flour showed no significant effects on anti/pro-oxidant parameters. However, lychee peel flour demonstrated notable effectiveness in reducing myocardial relaxation time in a T2DMOb model. These results suggest that while the antioxidant properties of lychee seed and peel flours were not significantly altered, lychee peel flour may have specific therapeutic potential to improve cardiac function in T2DMOb.
Acknowledgements
We are grateful to Priscilla Spadeto Altoé, RedOx e Inflammation Laboratory at UNESP-Botucatu (Brazil), UNIPEX (Unesp, Botucatu), and the LECTA, LHT and LHMI laboratories at the Federal University of Espírito Santo (Brazil) for their technical assistance. Also to Fundação de Amparo à Pesquisa e Inovação do Espírito Santo (FAPES, Brazil) and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Brazil) for their assistance.
Statement of Ethics
The animal experiments conform to internationally accepted standards and have been approved by the Animal Use Ethics Committee of the Federal University of Espírito Santo under protocol 24/2017; approved on August 4, 2017.
Author Contributions
PMC, ASL, and APLL conceived the design of the study. PMC, LMS, RRK, MFP, ASL, and APLL performed the experiments. PMC, ARM, STS, JCC, KCCS, ASL, and APLL interpreted and discussed the data and wrote the manuscript and statistical analysis. All authors revised the manuscript and the final version of the manuscript.
Funding
This work was supported by Fundação de Amparo à Pesquisa e Inovação do Espírito Santo – FAPES (grant number: 2022-VT4KM and 2022-BT5CX) and Higher Education Personnel Improvement Coordination – Capes.
Disclosure Statement
The authors have no conflicts of interest to declare.
References
| 1 | Bazmandegan G, Abbasifard M, Nadimi AE, Alinejad H, Kamiab Z. Cardiovascular risk factors in diabetic patients with and without metabolic syndrome: a study based on the Rafsanjan cohort study. Scientific Reports. 2023;13:559.
https://doi.org/10.1038/s41598-022-27208-5 |
| 2 | Diabetes [cited 2024 Jul 7] [Internet]. 2023 Available from: https://www.who.int/news-room/fact-sheets/detail/diabetes#:~:text=Diabetes%20is%20a%20chronic%20disease,hormone%20that%20regulates%20blood%20glucose.
|
| 3 | Eguchi K, Boden-Albala B, Jin Z, Rundek T, Sacco RL, Homma S, et al. Association between diabetes mellitus and left ventricular hypertrophy in a multiethnic population. The American journal of cardiology. 2008;101:1787-91.
https://doi.org/10.1016/j.amjcard.2008.02.082 |
| 4 | Lv Q, Si M, Yan Y, Luo F, Hu G, Wu H, et al. Effects of phenolic-rich litchi (Litchi chinensis Sonn.) pulp extracts on glucose consumption in human HepG2 cells. Journal of Functional Foods. 2014;7:621-9.
https://doi.org/10.1016/j.jff.2013.12.023 |
| 5 | Ferreira A, Matsubara L. Radicais livres: conceitos, doenças relacionadas, sistema de defesa e estresse oxidativo. Revista da associação médica brasileira. 1997;43:61-8.
https://doi.org/10.1590/S0104-42301997000100014 |
| 6 | Stefanović A, Kotur-Stevuljević J, Spasić S, Bogavac-Stanojević N, Bujisić N. The influence of obesity on the oxidative stress status and the concentration of leptin in type 2 diabetes mellitus patients. Diabetes research and clinical practice. 2008;79:156-63.
https://doi.org/10.1016/j.diabres.2007.07.019 |
| 7 | Punia S, Kumar M. Litchi (Litchi chinenis) seed: Nutritional profile, bioactivities, and its industrial applications. Trends in Food Science & Technology. 2021;108:58-70.
https://doi.org/10.1016/j.tifs.2020.12.005 |
| 8 | Wadhwa M, Bakshi M. Utilization of fruit and vegetable wastes as livestock feed and as substrates for generation of other value-added products. Rap Publication. 2013;4:67.
|
| 9 | Queiroz EdR, Abreu CMPd, Santos CMd, Simão AA. Composição química e fitoquímica das farinhas da casca e da semente de lichias (Litchi chinensis Sonn) cultivar'Bengal'. Ciência Rural. 2014;45:329-34.
https://doi.org/10.1590/0103-8478cr20140106 |
| 10 | Prasad KN, Yang B, Yang S, Chen Y, Zhao M, Ashraf M, et al. Identification of phenolic compounds and appraisal of antioxidant and antityrosinase activities from litchi (Litchi sinensis Sonn.) seeds. Food Chemistry. 2009;116:1-7.
https://doi.org/10.1016/j.foodchem.2009.01.079 |
| 11 | Man S, Ma J, Wang C, Li Y, Gao W, Lu F. Chemical composition and hypoglycaemic effect of polyphenol extracts from Litchi chinensis seeds. Journal of Functional Foods. 2016;22:313-24.
https://doi.org/10.1016/j.jff.2016.01.032 |
| 12 | Kilari EK, Putta S. Delayed progression of diabetic cataractogenesis and retinopathy by Litchi chinensis in STZ-induced diabetic rats. Cutaneous and Ocular Toxicology. 2017;36:52-9.
https://doi.org/10.3109/15569527.2016.1144610 |
| 13 | Sh R, Xu D, Pan Z, Gao Y, Gao Q. Two flavanone compounds from litchi (Litchi chinensis Sonn) seeds, one previously unreported, and appraisal of their a-glucosidase inhibitory activities. Food Chem. 2011;127:1760-3.
https://doi.org/10.1016/j.foodchem.2011.02.054 |
| 14 | Queiroz EdR, Abreu CMPd, Oliveira KdS, Ramos VdO, Fráguas RM. Bioactive phytochemicals and antioxidant activity in fresh and dried lychee fractions. Revista Ciência Agronômica. 2015;46:163-9.
https://doi.org/10.1590/S1806-66902015000100019 |
| 15 | Matias AM, Estevam WM, Coelho PM, Haese D, Kobi JBBS, Lima-Leopoldo AP, et al. Differential effects of high sugar, high lard or a combination of both on nutritional, hormonal and cardiovascular metabolic profiles of rodents. Nutrients. 2018;10:1071.
https://doi.org/10.3390/nu10081071 |
| 16 | Ferron AJT, Jacobsen BB, Sant'Ana PG, de Campos DHS, de Tomasi LC, Luvizotto RdAM, et al. Cardiac dysfunction induced by obesity is not related to β-adrenergic system impairment at the receptor-signalling pathway. PLoS One. 2015;10:e0138605.
https://doi.org/10.1371/journal.pone.0138605 |
| 17 | Jacobsen BB, Leopoldo APL, Cordeiro JP, Campos DHSd, Nascimento AFd, Sugizaki MM, et al. Perfis cardíaco, metabólico e molecular de ratos sedentários no momento inicial da obesidade. Arquivos Brasileiros de Cardiologia. 2017;109:432-9.
|
| 18 | Silva VL, Lima‐Leopoldo AP, Ferron AJ, Cordeiro JP, Freire PP, de Campos DH, et al. Moderate exercise training does not prevent the reduction in myocardial L‐type Ca2+ channels protein expression at obese rats. Physiological Reports. 2017;5:e13466.
https://doi.org/10.14814/phy2.13466 |
| 19 | Parveen K, Khan R, Siddiqui WA. Antidiabetic effects afforded by Terminalia arjuna in high fat-fed and streptozotocin-induced type 2 diabetic rats. Int J Diabetes Metab. 2011;19:23-33.
https://doi.org/10.1159/000497707 |
| 20 | Queiroz ER, Abreu CMD, Rocha DA, Sousa RVD, Fraguas RM, Braga MA, et al. Lychee (Litchi chinensis Sonn.) peel flour: effects on hepatoprotection and dyslipidemia induced by a hypercholesterolemic diet. Anais da Academia Brasileira de Ciências. 2017;90:267-81.
https://doi.org/10.1590/0001-3765201720150638 |
| 21 | Neves L, Alencar Sd, Carpes S. Determinação da atividade antioxidante e do teor de compostos fenólicos e flavonoides totais em amostras de pólen apícola de Apis mellifera. Brazilian Journal of Food Technology. 2009;7:107-10.
|
| 22 | Singleton VL, Rossi JA. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. American Journal of Enology and Viticulture. 1965;16:144-58.
https://doi.org/10.5344/ajev.1965.16.3.144 |
| 23 | Marques GS, Monteiro RPM, Leão WdF, Lyra MAM, Peixoto MS, Rolim-Neto PJ, et al. Avaliação de procedimentos para quantificação espectrofotométrica de flavonoides totais em folhas de Bauhinia forficata Link. Química Nova. 2012;35:517-22.
https://doi.org/10.1590/S0100-40422012000300014 |
| 24 | Favoretto S, Seabra D, Olivato M. Guia de eutanásia para animais de ensino e pesquisa. UNIFESP: São Paulo, Brazil. 2013:29-30.
|
| 25 | Taylor BA, Phillips SJ. Detection of obesity QTLs on mouse chromosomes 1 and 7 by selective DNA pooling. Genomics. 1996;34:389-98.
https://doi.org/10.1006/geno.1996.0302 |
| 26 | Matthews DR, Hosker JP, Rudenski AS, Naylor B, Treacher DF, Turner RC. Homeostasis model assessment: insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. diabetologia. 1985;28:412-9.
https://doi.org/10.1007/BF00280883 |
| 27 | Damiani APL, Caldas LC, Melo AB, Contreiro CDE, Estevam WM, Nogueira BV, et al. Resistance traning protocols promote strength increase without morphological changes. Revista Brasileira de Medicina do Esporte. 2020;26:253-7.
https://doi.org/10.1590/1517-869220202603209955 |
| 28 | Samarghandian S, Farkhondeh T, Samini F, Borji A. Protective effects of carvacrol against oxidative stress induced by chronic stress in rat's brain, liver, and kidney. Biochemistry research international. 2016;2016.
https://doi.org/10.1155/2016/2645237 |
| 29 | Uchiyama M, Mihara M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Analytical biochemistry. 1978;86:271-8.
https://doi.org/10.1016/0003-2697(78)90342-1 |
| 30 | Mesquita CS, Oliveira R, Bento F, Geraldo D, Rodrigues JV, Marcos JC. Simplified 2, 4-dinitrophenylhydrazine spectrophotometric assay for quantification of carbonyls in oxidized proteins. Analytical biochemistry. 2014;458:69-71.
https://doi.org/10.1016/j.ab.2014.04.034 |
| 31 | Marklund S, Marklund G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. European journal of biochemistry. 1974;47:469-74.
https://doi.org/10.1111/j.1432-1033.1974.tb03714.x |
| 32 | Pieper GM, Jordan M, Dondlinger LA, Adams MB, Roza AM. Peroxidative stress in diabetic blood vessels: reversal by pancreatic islet transplantation. Diabetes. 1995;44:884-9.
https://doi.org/10.2337/diab.44.8.884 |
| 33 | Lima-Leopoldo AP, Leopoldo AS, Da Silva DC, do Nascimento AF, de Campos DH, Luvizotto RA, et al. Long-term obesity promotes alterations in diastolic function induced by reduction of phospholamban phosphorylation at serine-16 without affecting calcium handling. Journal of applied physiology. 2014;117:669-78.
https://doi.org/10.1152/japplphysiol.00088.2014 |
| 34 | Jiang S, Ren D, Li J, Yuan G, Li H, Xu G, et al. Effects of compound K on hyperglycemia and insulin resistance in rats with type 2 diabetes mellitus. Fitoterapia. 2014;95:58-64.
https://doi.org/10.1016/j.fitote.2014.02.017 |
| 35 | Reuter TY. Diet-induced models for obesity and type 2 diabetes. Drug discovery today: disease models. 2007;4:3-8.
https://doi.org/10.1016/j.ddmod.2007.09.004 |
| 36 | Brito-Casillas Y, Melián C, Wägner AM. Study of the pathogenesis and treatment of diabetes mellitus through animal models. Endocrinología y Nutrición (English Edition). 2016;63:345-53.
https://doi.org/10.1016/j.endoen.2016.09.002 |
| 37 | Tang Y, Yu C, Wu J, Chen H, Zeng Y, Wang X, et al. Lychee seed extract protects against neuronal injury and improves cognitive function in rats with type II diabetes mellitus with cognitive impairment. International journal of molecular medicine. 2018;41:251-63.
https://doi.org/10.3892/ijmm.2017.3245 |
| 38 | Cheynier V, Tomas-Barberan FA, Yoshida K. Polyphenols: From plants to a variety of food and nonfood uses. Journal of Agricultural and Food Chemistry. 2015;63:7589-94.
https://doi.org/10.1021/acs.jafc.5b01173 |
| 39 | Padayachee A, Netzel G, Netzel M, Day L, Zabaras D, Mikkelsen D, et al. Binding of polyphenols to plant cell wall analogues-Part 2: Phenolic acids. Food chemistry. 2012;135:2287-92.
https://doi.org/10.1016/j.foodchem.2012.07.004 |
| 40 | Lian J-h, Xiang Y-q, Guo L, Hu W-r, Ji W, Gong B-q. The use of high-fat/carbohydrate diet-fed and streptozotocin-treated mice as a suitable animal model of type 2 diabetes mellitus. Scandinavian Journal of Laboratory Animal Science. 2007;34:22-9.
|
| 41 | Poteryaeva O, Usynin I. Dysfunctional high-density lipoproteins in diabetes mellitus. Problemy endokrinologii. 2022;68:69-77.
https://doi.org/10.14341/probl13118 |
| 42 | Ndrepepa G. High-density lipoprotein: a double-edged sword in cardiovascular physiology and pathophysiology. Journal of Laboratory and Precision Medicine. 2021;6.
https://doi.org/10.21037/jlpm-21-32 |
| 43 | Otocka-Kmiecik A, Mikhailidis DP, Nicholls SJ, Davidson M, Rysz J, Banach M. Dysfunctional HDL: a novel important diagnostic and therapeutic target in cardiovascular disease? Progress in lipid research. 2012;51:314-24.
https://doi.org/10.1016/j.plipres.2012.03.003 |
| 44 | Su D, Zhang R, Hou F, Chi J, Huang F, Yan S, et al. Lychee pulp phenolics ameliorate hepatic lipid accumulation by reducing miR-33 and miR-122 expression in mice fed a high-fat diet. Food & function. 2017;8:808-15.
https://doi.org/10.1039/C6FO01507G |
| 45 | Mhatre SV, Bhagit AA, Yadav RP. Proteinaceous pancreatic lipase inhibitor from the seed of Litchi chinensis. Food Technology and Biotechnology. 2019;57:113.
https://doi.org/10.17113/ftb.57.01.19.5909 |
| 46 | Caturano A, D'Angelo M, Mormone A, Russo V, Mollica MP, Salvatore T, et al. Oxidative stress in type 2 diabetes: impacts from pathogenesis to lifestyle modifications. Current Issues in Molecular Biology. 2023;45:6651-66.
https://doi.org/10.3390/cimb45080420 |
| 47 | Suzuki YJ. Oxidant-mediated protein amino acid conversion. Antioxidants. 2019;8:50.
https://doi.org/10.3390/antiox8020050 |
| 48 | Bollineni RC, Fedorova M, Blüher M, Hoffmann R. Carbonylated plasma proteins as potential biomarkers of obesity induced type 2 diabetes mellitus. Journal of proteome research. 2014;13:5081-93.
https://doi.org/10.1021/pr500324y |
| 49 | Dalle-Donne I, Rossi R, Giustarini D, Milzani A, Colombo R. Protein carbonyl groups as biomarkers of oxidative stress. Clinica chimica acta. 2003;329:23-38.
https://doi.org/10.1016/S0009-8981(03)00003-2 |
| 50 | Dworzański J, Strycharz-Dudziak M, Kliszczewska E, Kiełczykowska M, Dworzańska A, Drop B, et al. Glutathione peroxidase (GPx) and superoxide dismutase (SOD) activity in patients with diabetes mellitus type 2 infected with Epstein-Barr virus. Plos one. 2020;15:e0230374.
https://doi.org/10.1371/journal.pone.0230374 |
| 51 | Promyos N, Phienluphon PP, Wechjakwen N, Lainampetch J, Prangthip P, Kwanbunjan K. Inverse Correlation of Superoxide Dismutase and Catalase with Type 2 Diabetes among Rural Thais. Nutrients. 2023;15:2071.
https://doi.org/10.3390/nu15092071 |
| 52 | Yang Z, Zhang L, Liu J, Li D. Litchi Pericarp Extract Treats Type 2 Diabetes Mellitus by Regulating Oxidative Stress, Inflammatory Response, and Energy Metabolism. Antioxidants. 2024;13:495.
https://doi.org/10.3390/antiox13040495 |
| 53 | Emanuele S, Lauricella M, Calvaruso G, D'Anneo A, Giuliano M. Litchi chinensis as a functional food and a source of antitumor compounds: An overview and a description of biochemical pathways. Nutrients. 2017;9:992.
https://doi.org/10.3390/nu9090992 |
| 54 | Wang L, Lou G, Ma Z, Liu X. Chemical constituents with antioxidant activities from litchi (Litchi chinensis Sonn.) seeds. Food Chemistry. 2011;126:1081-7.
https://doi.org/10.1016/j.foodchem.2010.11.133 |
| 55 | Cardoso JC, Martins VVP, Madureira AR, Sales ST, Filetti FM, Corrêa CR, et al. A high-fat diet induces cardiac damage in obesity-resistant rodents with reduction in metabolic health. Cell Physiol Biochem. 2023;57:264-78.
https://doi.org/10.33594/000000642 |
| 56 | Gasparini PVF, Matias AM, Torezani-Sales S, Kobi J, Siqueira JS, Corrêa CR, et al. High-fat and combined high-fat and sucrose diets promote cardiac oxidative stress independent of Nox2 redox regulation and obesity in rats. Cell Physiol Biochem. 2021;55:618-34.
https://doi.org/10.33594/000000441 |
| 57 | Melo AB, Damiani APL, Coelho PM, de Assis ALEM, Nogueira BV, Ferreira LG, et al. Resistance training promotes reduction in Visceral Adiposity without improvements in Cardiomyocyte Contractility and Calcium handling in Obese Rats. International journal of medical sciences. 2020;17:1819.
https://doi.org/10.7150/ijms.42612 |
| 58 | Martins F, Campos DHS, Pagan LU, Martinez PF, Okoshi K, Okoshi MP, et al. High-fat diet promotes cardiac remodeling in an experimental model of obesity. Arquivos brasileiros de cardiologia. 2015;105:479-86.
https://doi.org/10.5935/abc.20150095 |
| 59 | Li Y, Liu Y, Liu S, Gao M, Wang W, Chen K, et al. Diabetic vascular diseases: molecular mechanisms and therapeutic strategies. Signal transduction and targeted therapy. 2023;8:152.
https://doi.org/10.1038/s41392-023-01400-z |
| 60 | Kishi S, Gidding SS, Reis JP, Colangelo LA, Venkatesh BA, Armstrong AC, et al. Association of insulin resistance and glycemic metabolic abnormalities with LV structure and function in middle age: the CARDIA study. JACC: Cardiovascular Imaging. 2017;10:105-14.
https://doi.org/10.1016/j.jcmg.2016.02.033 |
| 61 | Akhtar MS, Alavudeen SS, Raza A, Imam MT, Almalki ZS, Tabassum F, et al. Current understanding of structural and molecular changes in diabetic cardiomyopathy. Life Sciences. 2023:122087.
https://doi.org/10.1016/j.lfs.2023.122087 |
| 62 | Bers DM. Cardiac excitation-contraction coupling. Nature. 2002;415:198-205.
https://doi.org/10.1038/415198a |
| 63 | Zhang X, Qu L, Chen L, Chen C. Improvement of cardiomyocyte function by in vivo hexarelin treatment in streptozotocin‐induced diabetic rats. Physiological Reports. 2018;6:e13612.
https://doi.org/10.14814/phy2.13612 |
| 64 | Howarth F, Shafiullah M, Qureshi M. Chronic effects of type 2 diabetes mellitus on cardiac muscle contraction in the Goto‐Kakizaki rat. Experimental Physiology. 2007;92:1029-36.
https://doi.org/10.1113/expphysiol.2007.038703 |
| 65 | Jweied EE, McKinney RD, Walker LA, Brodsky I, Geha AS, Massad MG, et al. Depressed cardiac myofilament function in human diabetes mellitus. American Journal of Physiology-Heart and Circulatory Physiology. 2005;289:H2478-H83.
https://doi.org/10.1152/ajpheart.00638.2005 |
| 66 | Dong F, Yang X, Sreejayan N, Ren J. Chromium (D‐phenylalanine) 3 improves obesity‐induced cardiac contractile defect in ob/ob mice. Obesity. 2007;15:2699-711.
https://doi.org/10.1038/oby.2007.322 |
| 67 | Xu KY, Zweier JL, Becker LC. Hydroxyl radical inhibits sarcoplasmic reticulum Ca2+-ATPase function by direct attack on the ATP binding site. Circulation research. 1997;80:76-81.
https://doi.org/10.1161/01.RES.80.1.76 |
| 68 | Li S-Y, Yang X, Ceylan-Isik A, Du M, Sreejayan N, Ren J. Cardiac contractile dysfunction in Lep/Lep obesity is accompanied by NADPH oxidase activation, oxidative modification of sarco (endo) plasmic reticulum Ca 2+-ATPase and myosin heavy chain isozyme switch. Diabetologia. 2006;49:1434-46.
https://doi.org/10.1007/s00125-006-0229-0 |
| 69 | Hamouda NN, Qureshi MA, Alkaabi JM, Oz M, Howarth FC. Reduction in the Amplitude of Shortening and Ca 2+ Transient by Phlorizin and Quercetin-3-O-Glucoside in Ventricular Myocytes From Streptozotocin-Induced Diabetic Rats. Physiological research. 2016;65.
https://doi.org/10.33549/physiolres.933045 |
| 70 | Yao P, Gao Y, Simal-Gandara J, Farag MA, Chen W, Yao D, et al. Litchi (Litchi chinensis Sonn.): A comprehensive review of phytochemistry, medicinal properties, and product development. Food & Function. 2021;12:9527-48.
https://doi.org/10.1039/D1FO01148K |
| 71 | Bocchi L, Savi M, Naponelli V, Vilella R, Sgarbi G, Baracca A, et al. Long-term oral administration of theaphenon-e improves cardiomyocyte mechanics and calcium dynamics by affecting phospholamban phosphorylation and ATP production. Cellular Physiology and Biochemistry. 2018;47:1230-43.
https://doi.org/10.1159/000490219 |
| 72 | Zhao L, Wang K, Wang K, Zhu J, Hu Z. Nutrient components, health benefits, and safety of litchi (Litchi chinensis Sonn.): A review. Comprehensive Reviews in Food Science and Food Safety. 2020;19:2139-63.
https://doi.org/10.1111/1541-4337.12590 |
| 73 | Huang R, Shi Z, Chen L, Zhang Y, Li J, An Y. Rutin alleviates diabetic cardiomyopathy and improves cardiac function in diabetic ApoEknockout mice. European journal of pharmacology. 2017;814:151-60.
https://doi.org/10.1016/j.ejphar.2017.08.023 |
| 74 | Guimaraes JF, Muzio BP, Rosa CM, Nascimento AF, Sugizaki MM, Fernandes AA, et al. Rutin administration attenuates myocardial dysfunction in diabetic rats. Cardiovascular Diabetology. 2015;14:1-7.
https://doi.org/10.1186/s12933-015-0255-7 |