Impact of Micronutrients and Macronutrients on NK Cells Immunity
Keywords
Abstract
Natural killer (NK) cells are cytotoxic lymphocytes of the innate immune system. Along with their cardinal role in eliminating virally infected and cancerous cells, they are considered as a bridge between innate and adaptive immunity. Nutrition is linked to the effective immune response, and it is known that nutrition is among the environmental factors that influence the immune function and physiology. The function of nutrients, which are dissected into micronutrients (e.g. vitamins and minerals) and macronutrients (e.g. fat, protein and carbohydrates), is to maintain the metabolism and energy which are prime to fuel NK cells. In this review, we are going to recapitulate the recent findings and available data regarding the effect of common micro and macronutrients` examples on the NK cells development and function to provide an insight into diet–immune system crosstalk.
Introduction
Nutrients have long been known for sustaining growth, tissue development, providing energy for body metabolism, and shaping the immune system, which requires metabolic and nutritional cues for proper functioning [1, 2]. It is well documented that, nutrients deficiency or even insufficiency undermine the immune system [3]. The nutrients are considered chemical substances and are obtained ideally from a balanced diet. They are dissected into two broad categories: macronutrients (carbohydrates, proteins and fats) which are required in high amount and have central role in energy provision [4] and micronutrients (vitamins and minerals) that are needed in low amount and their primary role is metabolism and maintaining tissue function [5, 6].
Vitamins are organic molecules and are categorized into; water-soluble (vitamins B1, B2, B3, B5, B6, B7, B9, B12, and C) and fat-soluble (vitamins A, E, D, and K) [7]. Minerals are inorganic micronutrients and could be classified based on their daily requirement into macrominerals and required in greater amount (calcium, phosphorous, magnesium, sodium, potassium, and chloride) or microminerals which are required in smaller amount ( iron, copper, zinc, selenium, and iodine) [8].
NK cells are innate lymphoid cells and able to eliminate virally infected and transformed cells by means of cytotoxicity and cytokines secretion. NK cells have a unique role in shaping the innate and adaptive immunity and they have a defensive alliance with other immune cells upon antigen encounter [9-11]. Previous studies showed that NK cells engage to concrete metabolic responses that are linked to their cell effector functions [2, 12].
Stemming from the fact that nutrients are pivotal to fulfil the metabolism within immune cells, including NK cells, and subsequently affecting their development and activity, we aimed in this review to summarize the recent and existing data concerning the effects of macronutrients and micronutrients on NK cells` metabolism, development and activity in the context of experimental and clinical studies to provide better understanding for the intricate and dynamic link between nutrients and NK cells in normal and aberrant conditions Table 1 &2.
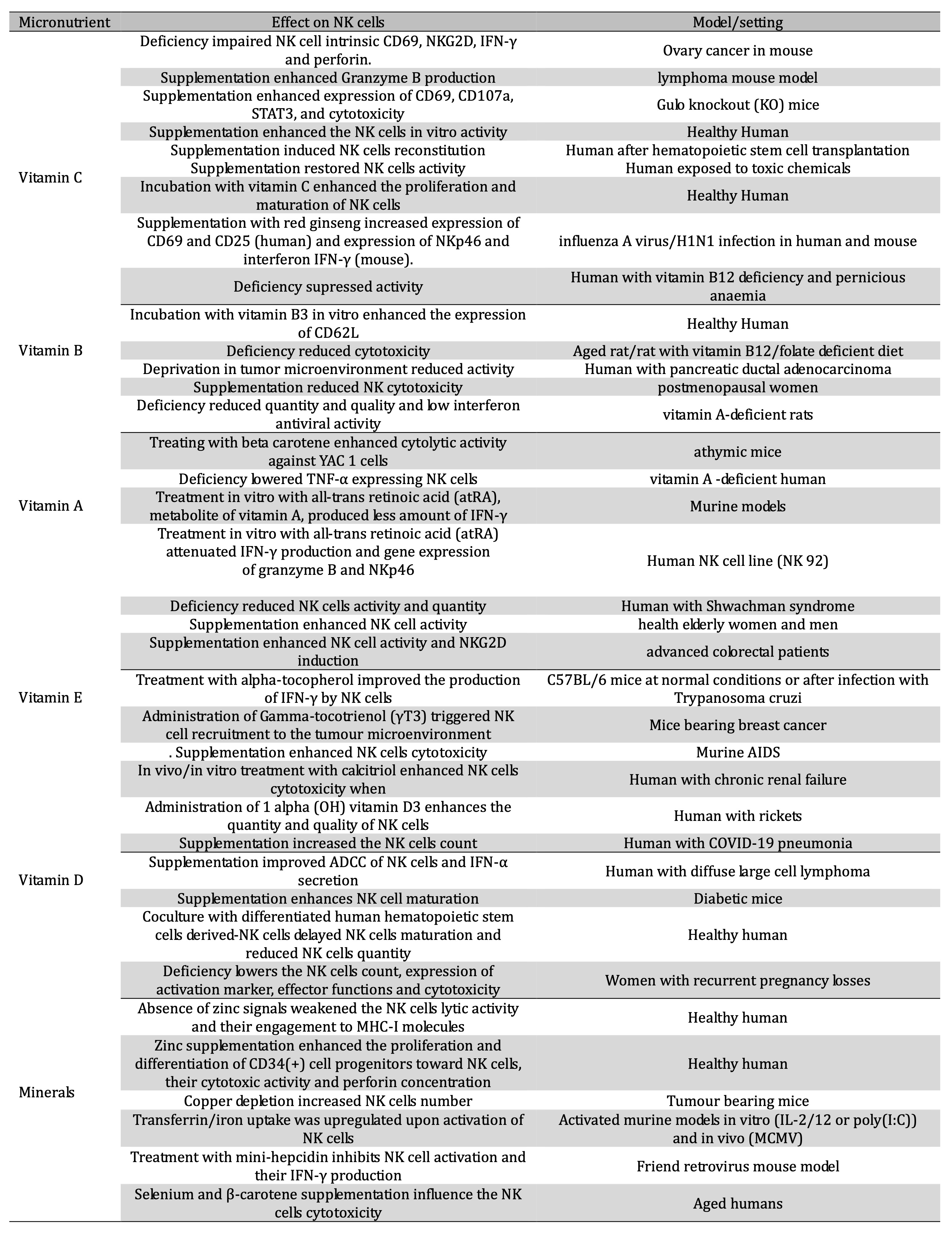
Table 1: Impact of micronutrients on NK cells
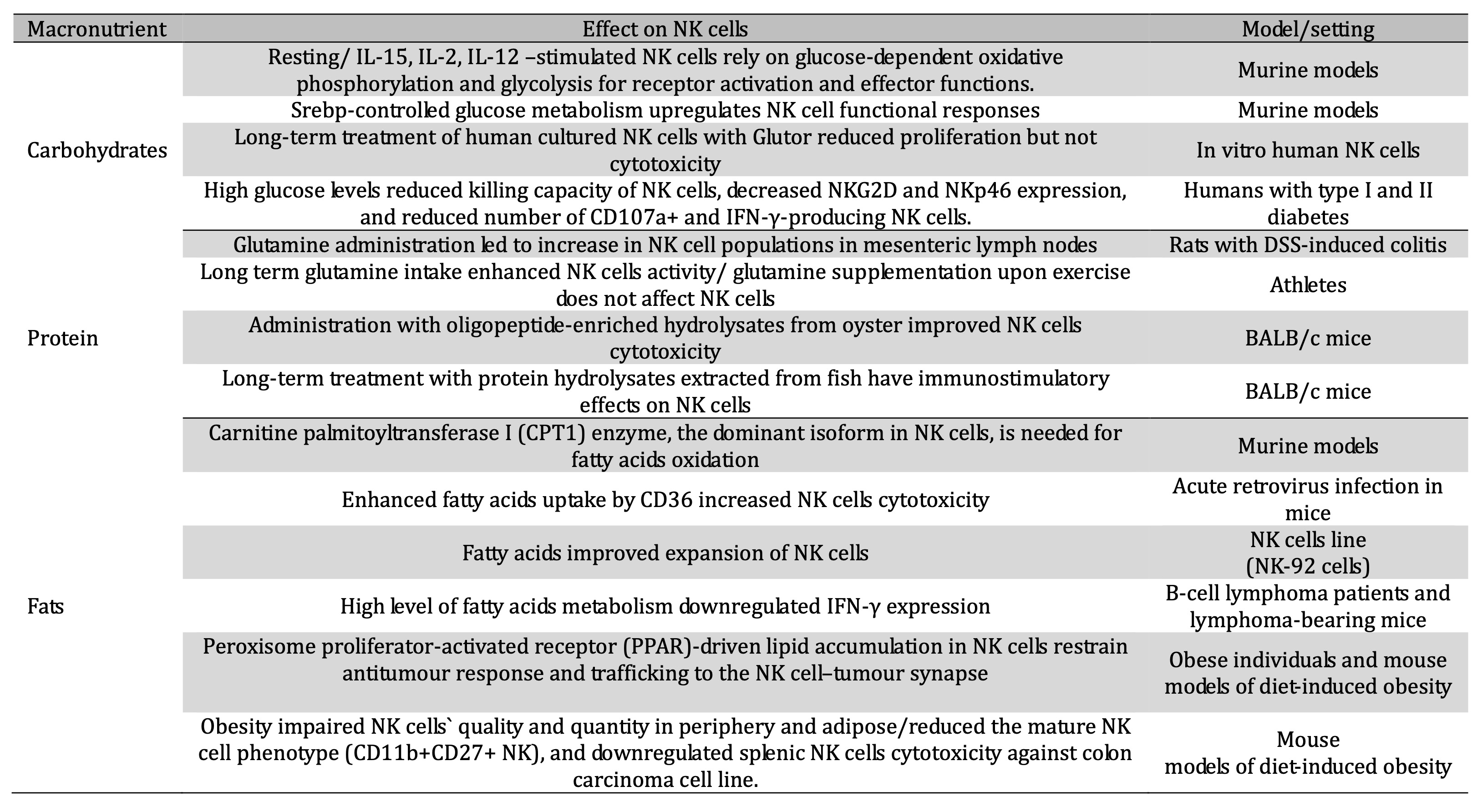
Table 2: Impact of macronutrients on NK cells
Effect of Micronutrients
Vitamin C
Vitamin C, also known as ascorbic acid
(AA), is water soluble vitamin, and is regarded as an antioxidant
with many biological and immunological functions [13]. Humans are
unable to synthesize vitamin C as they lack L-gulono-gamma-lactone
oxidase, the enzyme catalysing the terminal step in L-ascorbic acid
biosynthesis [14], and thereby it must be taken by diet or as
supplements [15].
It has long been claimed that vitamin C
has vast roles in the context of normal functioning of the immune
system, and it exerts a wide spectrum of favorable effects on the
innate and adaptive cellular compartments of the immune system
including NK cells. The role of ascorbic acid in regulating the
function of NK cells was reported in many compelling experimental and
clinical studies.
For example, Kim et al. showed that
ascorbic acid depleted mice succumb to ovarian cancer compared to the
control group. NK cell-intrinsic CD69 and NKG2D were greatly impaired
in ascorbic acid deficient mice, also IFN-γ and perforin secretion
was downregulated, substantiating the role of ascorbic acid in
maintaining the NK cells cytotoxicity against cancer [16]. In
parallel, using high dose ascorbic acid treatment in combination with
anti-PDL1 therapy improved the Granzyme B production in NK cells in
the context of lymphoma mouse model [17].
Another series of studies conducted on
human demonstrated the importance of vitamin C in modulating the NK
cells activity. For example, administration of human with ascorbic
acid supplement enhanced the NK cells activity 8 hours post treatment
when tested with 4-hr.-51Cr-release
assay using K562 tumor cells as targets [18]. Furthermore, vitamin C
supplementation to patients after hematopoietic stem cell
transplantation induced NK cells reconstitution [19].
Further findings regarding NK cell
activation by AA were also evidenced by Huijskens and colleagues.
Incubation of mature NK cells isolated from peripheral blood
mononuclear cells with vitamin C enhanced the proliferation of NK
cells without effect on NK cells cytotoxicity. Moreover, ascorbic
acid enhanced the NK cell maturation from early T/NK-cell progenitors
[20].
In the context of viral infections,
vitamin C plays a substantial role in the NK cell biology. For
instance, synergistic administration of vitamin C with red ginseng
during influenza A virus/H1N1 infection enhanced the NK cells
activity in both human and murine models as demonstrated by increased
expression of CD69 and CD25 on human NK cells after synergistic
treatment for 48 hours and improved expression of NKp46, a natural
cytotoxic receptor of NK cells and IFN-γ generation and thereby
amelioration of virus-induced inflammation [21].
B vitamins
B group vitamins are among water soluble
vitamins that have a manifold of cellular functions including
regulatory and metabolic roles [22]. The vitamins that make up B
complex are thiamine (vitamin B1), riboflavin (vitamin B2), niacin
(vitamin B3), pantothenic acid (vitamin B5), pyridoxine (vitamin B6),
biotin (B7), folate (B9), and cobalamin (vitamin B12). Most vitamins
B should be taken by diet as human can not synthesize it. Another
potent source of vitamins B are bacteria inhabiting the gut [22]. It
is noteworthy that vitamin B1 (thiamin) was the first vitamin
identified [23]. B vitamins are linked to the immune system with a
myriad of immunomodulatory roles.
A compelling body of research studies
reported the crosstalk between B vitamins and immune regulation
mediated by NK cells. A study on patients with vitamin B12 deficiency
showed that their NK cells have supressed activity, and the activity
was enhanced after methyl-B12 treatment. Nevertheless, there was no
improvement on NK cell activity after methyl-B12 supplementation in
vitamin B12 sufficient individuals [24]. By the same token, patients
with pernicious anaemia, a type of vitamin B12 deficiency, have
reduced NK cell activity, which was restored after cyanocobalamin
treatment [25]. Moreover, addition
of Nicotinamide, a form of vitamin B3, enhanced the expression of
CD62L on in vitro
expanded NK cells and thereby improved
their recruitment to bone marrow and lymph nodes [26].
In parallel, splenic NK cells harvested
from aged rat with vitamin B12 deficient
diet exhibited reduction in NK killing ability for YAC 1 cells,
murine melanoma cell line sensitive to NK cell mediated killing [27].
Further, severe folate deficiency impairs NK cells cytotoxicity in
rats [28].
In contrast, a clinical study on
individuals with supraphysiological levels of folate showed that high
levels of folate are not associated with improved NK cell activities
[29]. Another similar in vitro
study showed that, NK cells cytotoxicity
was not impacted by high levels of folate [30]. In accordance, the
quality and quantity of NK cells was not associated with the
concentration of folate in healthy individuals [31]. A study
conducted on postmenopausal women showed that utilizing a folate-rich
diet and folic acid supplements led to reduced NK cytotoxicity
compared with those consuming a low-folate diet and no supplements
[32].
Vitamin A
Vitamin A is indispensable fat-soluble
micronutrient that is stored primarily in liver after its digestion
and absorption in the small intestine [33]. Upon inflammatory
process, recruited NK cells are exposed to vitamin A-enriched
microenvironment [34-36]. Earlier study showed that vitamin
A-deficient rats have fewer number of splenic NK cell [37] and
lower cytotoxicity among splenic [38], and peripheral NK cells [39].
Supplementation of these rats with retinoic acid, a metabolite of
vitamin A, restored the quantity and quality of NK cells to normal
levels [37]. Along with retinoic acid, the impact of beta carotene on
NK cell activation was elucidated. For instance, treating NK cells
from athymic mice with beta carotene led to enhanced cytolytic
activity against YAC1 cells [40, 41]. In consistent, human study on
Ghanaians population demonstrated that vitamin A -deficient group was
linked to lower functional NK cells as mirrored by reduced TNF-α
expressing NK cells [42]. Another study conducted on human NK cell
line (NK 92) which are expressing receptors for All-trans retinoic
acid (ATRA) showed that treatment NK 92 cell line with ATRA
attenuated IFN-γ production and gene expression of granzyme
B and NKp46, suggesting reduced cytotoxicity [43].
Vitamin E
Vitamin E is a powerful fat soluble
antioxidant, and it exists in high concentration in immune cells
compared to other cells [44]. Vitamin E could be stored in body, so
vitamin E does not have to be consumed every day. Vitamin E
encompasses four tocopherols (α-, β-, γ-, and δ-tocopherols) and
four tocotrienols (α-, β-, γ-, and δ-tocotrienols) and are
present in food, and only α-tocopherol meets the human vitamin E
need [45, 46]. It has a potential immune modulatory role, due to its
role in protection against oxidation of polyunsaturated fatty acids
which are major components of plasma membrane for immune cells [44].
The function of vitamin E in regulating immune system was
investigated in different mice and human studies during normal and
disease conditions, and these studies showed that vitamin E appears
to be linked with the NK cell functionality.
Examination of NK cells from a Japanese
boy suffering from Shwachman syndrome associated with severe vitamin
E deficiency showed that NK cells activity and the number of CD56dim
CD16+
NK cells were
diminished. In the same case report, eight-weeks alpha-tocopherol
supplementation reversed the cytotoxic NK cells abnormality in terms
of quality and quantity [47, 48]. In consistent, dietary
supplementation of vitamin E (200mg daily) was reported to enhance NK
cell functions in healthy elderly women and men [49]. Another similar
study conducted on elderly women showed that NK cell activity was
positively corelated with vitamin E concentration [31]. Further,
short term supplementation of vitamin E to advanced colorectal
patients enhanced NK cell activity in PBMC when cocultured with 51Cr
labeled K562, partly due to marginal induction of NKG2D expression,
nevertheless, the ability of NK cells to generate perforin and IFN-γ
was unaffected by vitamin E administration [50].
Studies on animals also supported the
positive impact of vitamin E on NK cells. A recent study showed that
treatment of C57BL/6 mice with alpha-tocopherol improved the
production of IFN-γ by NK cells at normal conditions and after
infection with Trypanosoma
cruzi which resulted in lower
parasitemia, attenuated tissue destruction and fibrosis and thereby
less mortality [51]. Another analogue of vitamin E was also studied.
For instance, Gamma‐tocotrienol
(γT3) administration in mice bearing breast cancer triggered NK cell
recruitment to the tumour microenvironment resulting in reduced
expression of TLR proteins in the tumour microenvironment and hence
suppressing the tumour growth [52]. In addition, the effect of
vitamin E on NK cells was also analysed in the context of murine
AIDS. The suppressed NK cells cytotoxicity during retrovirus
infection in mice was enhanced after vitamin E supplementation [53].
Vitamin D
Vitamin D is an essential micronutrient
for bone health and mineral metabolism including calcium [54]. It is
a secosteroid hormone and exists in two major forms: ergocalciferol
(D2) and cholecalciferol (D3) [55, 56]. Vitamin D2 is created by
plants and fungi via UV irradiation of ergosterol, a
steroid found in the plants and fungi. Cholecalciferol (vitamin D3)
is synthesized via the UV irradiation of 7-dehydrocholesterol.
Generally, vitamin D requirement is sourced from sunlight exposure,
or ingestion which include habitual dietary intake, fortified foods
or as supplementation [57].
1α,25-dihydroxyvitamin D3 (1,
25(OH)2D3),
also known as calcitriol, is regarded as the active form of vitamin D
and exerts immunoregulatory functions via vitamin D receptor (VDR)
which are expressed by numerous immune cells including NK cells [58,
59], substantiating the role of vitamin D on regulating the NK cells
biology. More specifically, NK cells require vitamin D receptors for
development [60].
A clinical study on individuals of
chronic renal failure showed that these patients have reduced
secretion of calcitriol and subsequent less NK cell cytotoxicity. In
vivo treatment with calcitriol
or in vitro
treatment of PBMC retrieved from patients
and controls resulted in enhanced NK cells cytotoxicity when
cocultured with chromium labelled target cells [61]. Another study on
patients with rickets demonstrated that administration of 1 alpha
(OH) vitamin D3 enhances the quantity and quality of NK cells [62]. A
cohort study on vitamin D insufficient and deficient COVID-19
pneumonia patients showed that individuals with deficient vitamin D
have lower NK cells count [63]. Subsequent study performed on
patients with severe and critical COVID 19 patients admitted to ICU,
revealed that vitamin D3 supplementation remarkably increased the NK
cells count compared to placebo [64].
Vitamin D was reported to improve the
outcome of immuno-chemotherapy with rituximab in patients suffering
from diffuse large cell lymphoma by increasing the ADCC of NK cells,
due to NK cell intrinsic IFN-α secretion and thereby increased
killing ability against rituximab-coated target cell which is further
harnessed by production of IL-2 and IL-17 [65].
Furthermore, the role of vitamin D in
modulating NK cells response was also delineated in the murine models
during diabetic conditions and obesity where the immune system is
dysfunctional [66]. A recent study showed that, NK cells from
diabetic mice exhibited diminished activity and maturation. Dietary
supplementation of vitamin D3 in diabetic mice enhanced NK cell
maturation as mirrored by increased percentage of CD11b
single-positive NK and NKG2D positive cells, and the NK cell mature
phenotype is linked to better activity. In addition, vitamin D3
administration enhanced the ability of NK cells to secrete IL-12 and
to express Bcl2 and Tbx21
which are key transcriptional factors for
NK cells maturation [67].
Even though the findings regarding the
positive impact of vitamin D on NK cells quality and quantity are
consistent, a substantial body of evidence showed the negative
influence of vitamin D on NK cells. For instance, coculture of 1,
25(OH)2D3
with in
vitro differentiated human
hematopoietic stem cells derived-NK cells resulted in delay in NK
cells maturation, reduced frequency and number of NK cells [68].
Nearly 50 % of women with recurrent
pregnancy losses have deficiency in vitamin D levels, lower NK cells
count, and downregulated cytotoxicity compared to the women with
sufficient vitamin D levels [69]. Another similar study showed that,
addition of 1, 25(OH)2D3 to NK cells obtained from PBMC harvested
from women with recurrent pregnancy losses led to NK cells
cytotoxicity suppression, CD107a, NCRs and CD69 expression
downregulation, reduced production of IFN-γ or TNF-α and increased
expression of NK cells inhibitory receptors [70].
Minerals
Originally, nutrient minerals are
regarded as metals and are distinct inorganic compounds. Minerals
have a myriad of physiological functions, and they are vital
components of different bioactive molecules such as hormones and
enzymes. Based on their daily requirement, minerals could be
categorised into; macrominerals, secondary and microminerals (trace
minerals) [71, 72]. Certain minerals are crucial for efficient immune
system functioning and NK cells biology [73].
Zinc deficiency was reported to
negatively influence the NK cells activity. For instance, the lytic
activity and engagement to the major histocompatibility complex class
I (MHC-I) on target cells are weakened when the zinc signals are
absent [74-76]. Furthermore, zinc supplementation enhanced the
proliferation and differentiation of CD34(+) cell progenitors toward
NK cells, their cytotoxic activity and perforin concentrations in
healthy human [77-79].
Copper was reported to regulate the
immunological synapse across many cancers. It was shown that, copper
depletion using chelators, tetraethylenepentamine (TEPA),
downregulated phosphorylation of intracellular signalling molecules
(STAT3 and EGFR) and induced ubiquitin-mediated degradation of PD-L1.
Also, Copper-chelating drugs increased NK cells and hampered tumour
growth in murine models [80]
NK cells activity is also dependent on
iron haemostasis. For example, CD71, transferrin receptor needed for
transferrin/iron uptake, was upregulated upon activation of NK cells
by poly(I:C) in vivo [81]
or stimulation by IL-2/12
in vitro [12]
and after activation with MCMV or friend
retroviral infection [82], indicating the necessity of iron to NK
cells activity [82]. In the same context, mice treatment with
mini-hepcidin, synthetic polypeptide that reduce iron level, inhibits
NK cell activation and their IFN-γ production during friend
retroviral infection [82].
Selenium is another essential
micronutrient with antioxidant properties [83], and it has been
reported that serum selenium concentration is positively correlated
with increased NK cells numbers but not the function in elderly
individuals [31] whereas selenium
supplementation in the same study influences the NK cells
cytotoxicity rather than the NK cells number. Contrasting study
showed that, the effect of selenium on NK cells is effective only
with β-carotene administration and is dependent on the period of
supplementation [84].
Effect of Macronutrients in NK cells
Carbohydrates
Carbohydrates are carbon containing
macromolecules and are essential for metabolism and energy production
to fuel cells for proper function and development [85]. Carbohydrates
exist in numerous forms including monosaccharides, disaccharides and
polysaccharides. Here we are going to present the findings and
existing data regarding the effect of glucose, the simplest form of
carbohydrates, on NK cells activity.
The dependence of NK cells on glucose is
essential for their activation and to exerts their functions and
certain NK cells rely on metabolic cues and have specific metabolic
requirements for IFN-γ and granzyme B production [86, 87]. In more
details, resting NK cells require glucose-dependent oxidative
phosphorylation (generation of ATP in the presence of oxygen), where
as expansion in the presence of IL-15 rely on glycolysis (metabolic
breakdown of glucose) [88, 89]. Releasing IFN-γ is also found to be
dependent on glycolysis and oxidative phosphorylation upon short term
receptor activation. In contrast, IFN-γ production induced by
activation of (IL-12 + IL-18) does not rely on both metabolic
pathways. Similarly, receptor activation depends minimally on these
metabolic pathways after priming with high-dose IL-15 [88]. Others
have demonstrated that, glycolysis is required for efficient
generation of IFN-γ and granzyme B in activated NK cells upon
expansion in IL-15 and stimulation with IL-2 + IL-12. The mammalian
target of rapamycin (TORC1) activity is shown to be crucial for
achieving this elevated glycolytic state [81].
Furthermore, glucose is not only
essential for maintaining NK cells` effecter functions but also to
cater various biosynthetic pathways. It was reported that glycolysis
and oxidative phosphorylation rates were mediated by sterol
regulatory element-binding protein, Srebp, which is crucial to
upregulate the proliferation and effector function of NK cells [90].
Moreover, long-term treatment of human
cultured NK cells with Glutor, an inhibitor of glucose transporters,
resulted in impeding NK cells proliferation due to decreased
glycolysis, with less effect on cytotoxicity [91].
The conditions where the glucose levels
are impaired have an effect on NK cells activity. Kim et
al, showed that patients with
type 2 diabetes have lower NK cells activity [92]. Consistently,
analysis of NK cell subsets in patients with long standing DM type 2
have a remarkable decrease in NK cells expressing NKG2D and NKp46
along with reduced killing capacity [93]. Also, untreated type 1 DM
children and adults have remarkable decrease count for different NK
cells subsets and have significant low number of CD107a+ and
IFN-γ-producing NK cells [94].
Proteins
Proteins are essential macronutrients.
The amino acids constituents of proteins orchestrate the immune
response by activation of the immune cells including NK cells,
assisting the lymphocyte proliferation and generating soluble
mediators such as; antibodies and cytokines [95]. Early studies
suggested that, amino acids are key immune regulators and energy
substrates for the immune cells [96]. Considering the instrumental
role of amino acids in shaping the immune response, it is not
surprising that protein malnutrition is linked to immune deficiency
[97].
The involvement of amino acids in
regulating the NK cell’s function has been implicated in numerous
evidence. A study on rats with DSS-induced colitis exhibited that
glutamine administration led to increase in NK cell populations in
mesenteric lymph nodes [98]. Another study conducted on human showed
that, athletes who took glutamine for three weeks with heavy load
training have enhanced NK cells activity relative to the control
group [99]. These findings were inconsistent with another study which
showed that NK cells were unchanged after glutamine supplementation
upon exercise [100].
Some food proteins mediate generation of
bioactive peptides (protein hydrolysates). They are encrypted in the
sequence of their parent proteins, and have a greater array of
bioactivities including immunoregulatory roles [101, 102]. The
immunomodulatory role of the protein hydrolysates in enhancing the NK
cells activity has been addressed in different studies. Oral
administration of BALB/c mice with oligopeptide-enriched hydrolysates
from oyster improved NK cells cytotoxicity [103, 104]. Further,
Long-term treatment of BALB/c mice with protein hydrolysates
extracted from fish have immunostimulatory effects on NK cells [105,
106].
Fats
Fats are central macronutrients and made
up of blocks called fatty acids, which are major components of plasma
membrane. Fatty acids are essential for energy production, and
cellular signalling. Furthermore, fatty acids are regarded as
precursors for the synthesis of lipid compounds needed for the
regulation of immune responses and inflammatory pathways [1, 107].
It has been reported that fatty acids
oxidation constitutes the major source of energy for NK cells in the
tumour microenvironment [108], where the glucose are often deprived
and the fatty acids are abundant [109, 110]. The fatty acids
oxidation relies mainly on enzyme carnitine palmitoyltransferase I
(CPT1), of which CPT1A is the dominant isoform in NK cells. Ablation
of CPT1A restrains long-chain fatty acid access into the mitochondria
[111]. Sheppard et al. found that active NK cells due to infection or
tumour enhanced fatty acid uptake and CPT1A expression by maintaining
NK cells` mitochondrial functions. The dependence of NK cells on
fatty acids oxidation was found to be more pronounced in active NK
cell mediated by receptor ligation compared to cytokine-driven NK
cells [108].A recent study exhibited that, intermittent fasting
improved the fatty acids oxidation which is mediated by increased
expression of the enzyme CPT1A, resulting in enhanced NK cells
survival and effector functions and thereby better NK cells anti-
tumour activity [112]. Also, the activated NK cells increased the
expression of CD36, fatty acid uptake receptor, and subsequent
enhanced fatty acids uptake, culminating in enhanced cytotoxicity
against viral infections [113]. Furthermore, fatty acids were found
to improve expansion of NK cell line (NK-92 cells) in
vitro by enhancing oxidative
phosphorylation and energy metabolism of NK-92 cells [114].
In contrast, NK cells retrieved from
B-cell lymphoma patients and from lymphoma-bearing mice had high
level of fatty acids metabolism, impaired mitochondrial function and
downregulated IFN-γ expression. This reduced functionality of NK
cells was attributed to excess fatty acids in the lymphoma
environment which impair the NK cell’s function. NK cells tend to
display high expression of the transcriptional regulator peroxisome
activator receptor-γ (PPAR-γ) which support their metabolism and
function, as a kind of adaptation to sustain their function in the
lymphoma environment [115]. In contrast, obesity which is a leading
cause for certain types of cancer stimulates peroxisome
proliferator-activated receptor (PPAR)-driven lipid accumulation in
NK cells and restrains anti-tumour response and trafficking to the NK
cell–tumour synapse in in
vivo and in
vitro settings [116]. Previous
studies on animals also exhibited the effect of obesity on the
impairment of NK cells` quality and quantity in periphery and adipose
tissues [117-119]. Spielmann et
al. reported that mice fed
with high fat diet exhibited reduced percentage of total NK cells and
the mature NK cell phenotype (CD11b+CD27+ NK).
In the same study, the splenic NK cells cytotoxicity was
dysfunctional against colon carcinoma cell line [120].
Mechanistic approach for NK response to nutrients
Different mechanisms drive the NK response toward the nutrients in direct and indirect manner via intracellular signaling molecules, metabolic regulators, receptor/ligand axis, proinflammatory cytokines, and via intermediate molecules.
Effect via intracellular signaling molecules
Some nutrients modulate NK cells response
directly via affecting intracellular signaling molecules which are
required to drive NK cell activation, such as STAT3 and protein
kinase C (PKC). For example, it
has been reported that the splenocytes harvested from Gulo knockout
(KO) mice supplemented with Aptamin
C, combination of vitamin C and aptamer, showed enhanced expression
of CD69 and CD107a and increased STAT3 activation [121]. Further, the
ability of NK cells to kill cocultured YAC-1 cells was improved in
mice received Aptamin C. It is noteworthy that using aptamer with
vitamin C enhance its stability and persistence in the body [122].
Exposure to toxic chemicals impaired NK cells functionality, which
was restored by vitamin C supplementation, and this was attributed to
the PKC [123]. Zinc has a role in signal transduction which is a
pillar in the context of the activation and inhibitory receptors
[77]. On contrary, the negative effect of 1, 25(OH)2D3 on NK cells
from women with recurrent pregnancy losses was due to the fact that
1, 25(OH)2D3 downregulates TLR4 [124], and the TLR4 is essential for
activation of human and murine NK cells [125].
Effect via metabolic regulators
The metabolic regulators such as mTORC1
and cMyc are needed for glycolysis and upregulation the
proinflammatory signature for NK cells in different settings. Recent
study demonstrated that murine NK cells treated in
vitro with all-trans retinoic
acid (atRA), metabolite of vitamin A, produced less amount of
IFN-γ and have reduced mitochondria mass and production of reactive
oxygen species. NK cells treated with atRA failed to engage mTORC1
and cMyc [36]. Another compelling study showed that glutamine
transport through SLC7A5, l-amino acid transporter in activated NK
cells, regulates cMyc protein expression in NK cells, and this
expression reported to be essential to shape NK cells growth and to
exert their effector functions, culminating in improved anti-tumour
activity [126].
Effect via receptor/ligand axis
Nutrients might influence via the
receptor/ligand axis. Zinc was reported to be involved in the
polymerization of Inhibitory killer-cell Ig-like receptor (KIR) and
it induces KIR assembly into filaments at the plasma membrane of NK
cells [76]. Moreover, reduced expression of NKp30/p46 on NK cells in
the context of type I diabetes [127], is explained by a decrease in
the synthesis and an increase in the degradation of heparin sulfate
proteoglycans [128]; these molecules are regarded as ligands for both
NKp46 and NKp30 [129]. In addition, folate, a prime cofactor in DNA
methylation process, could regulate the expression of KIRs and
subsequently modify NK cell activity [130].
Effect via proinflammatory cytokines
Other than interfering with the
signalling pathways. Nutrients can affect NK cells indirectly via
proinflammatory cytokines such as IL-2. Rodacki et
al. showed that patients with
long standing type 1 diabetes have diminished NK cells activation
compared to healthy controls or recent-onset patients, as mirrored by
downregulation of NKp30/p46 and of IFN-γ and perforin mRNA. This
could be attributed to the impaired secretion of IL-2, which induce
the NK cell activation [127]. The positive effect of calcitriol on NK
cells activity could be due to the direct impact of the calcitriol on
NK cell effector functions or indirectly via modulating other immune
cells as monocytes or lymphocytes which affect NK cells via a myriad
of cytokines (e.g.IL-1) [131]. Upon the treatment of in
vitro differentiated human
hematopoietic stem cells derived-NK cells with 1, 25(OH)2D3, NK cells
exhibited delay in NK cells maturation, and reduced quantity due to
antiproliferative effect of 1, 25(OH)2D3 on NK cells and suppression
of IL-2 production from helper T cells, which is essential cytokine
for NK cells activation [68, 132].
Effect via metabolic programming
Shaping the metabolic programming of NK
cells is another approach of how nutrients modulate NK cells. A
recent study exhibited that the growth of pancreatic ductal
adenocarcinoma requires high amount for vitamin B6 culminating
in the deprivation of vitamin B6 in the tumour microenvironment.
Scarcity of vitamin B6 leads to reduction in NK cells activity as the
vitamin B6 is critically needed for intracellular glycogen breakdown
in NK, which is necessary for NK cells effector functions [133].
Oxidative stress and endoplasmic reticulum stress are known to be
induced by high glucose levels [134], which possibly modulate the NK
cells activity including the NKG2D expression [93, 135]. Strikingly,
this inhibitory effect of glucose was reversed when treatment of NK
cells from diabetic individuals with IL-15, key cytokine for NK cells
maturation and NKG2D expression [93, 136].
Effect through intermediate molecules
Nutrients could affect NK cells
indirectly via intermediate molecules, and this is observed through
the crosstalk between nitric oxide and vitamin E in modulating NK
cells effector functions It is reported that vitamin E affects nitric
oxide levels [137], and the NK cells- intrinsic nitric oxide, which
is produced in low amounts, could positively improve NK cells
activity [138, 139]. In turn, NO could be detrimental as observed
from a study conducted by Stiff et.al,
in which they found that higher amount of nitric oxide produced by
myeloid-derived suppressor cells impairs NK cells functionality in
melanoma patients [140]. Another example is the indirect effect of
vitamin D on NK cells through calcium. One study showed that NK cell
effector functions via exocytosis of lytic granules is mediated by
calcium levels which is determined by calcitriol [141].
Concluding remarks
Macro- and micro- nutrients have a
substantial impact on NK cells by affecting their metabolism, quality
and quantity in humans and preclinical models in the context of
normal and aberrant conditions. While the favourable effect of
nutrients on NK cells was consistent in a substantial body of
studies, some contrasting reports demonstrated the negative influence
of nutrients on NK cells biology. This review provides insights into
the link between nutritional Immunology and immunometabolism field.
As NK cells are critically involved in the defence mechanism against
viruses and transformed cells and since the NK cells-based therapy is
pillar in certain cancers, understanding the crosstalk between
nutrients and NK cells is fundamental to establish feeding regimen
for disease prevention and patient care.
Future perspectives
In this review, we shed a light on the
effect of nutrients on NK cells biology. In one hand, NK cells
require high demands of nutrients as fuel to function properly in
different conditions. On other hand, the nutrients could be
repercussion for NK cells. Based on the nutrient nature and setting,
the nutrient could be used as an adjuvant substance and
non-pharmacological intervention to boost NK cells, or could be
deprived to enhance NK cells response, this would be more important
in the rapidly evolving cancer field. As we explored the current
knowledge of dietary impact on NK cells, we posit that further
investigations in the field of nutritional immunology needs a merit.
Acknowledgements
Author Contributions
Conceptualization, T.A.H; resources,
T.A.H.; writing—original draft preparation, T.A.H.; writing—review
and editing, T.A.H. All authors have read and agreed to the published
version of the manuscript.
Disclosure Statement
The authors have no conflicts of interest to declare.
References
| 1 | Tourkochristou E, Triantos C and Mouzaki A: The Influence of Nutritional Factors on Immunological Outcomes. Front Immunol 2021;12:665968.
https://doi.org/10.3389/fimmu.2021.665968 |
| 2 | Zhang P, Watari K and Karin M: Innate immune cells link dietary cues to normal and abnormal metabolic regulation. Nature Immunology 2025;26:29-41.
https://doi.org/10.1038/s41590-024-02037-y |
| 3 | Wu D, Lewis ED, Pae M and Meydani SN: Nutritional Modulation of Immune Function: Analysis of Evidence, Mechanisms, and Clinical Relevance. Front Immunol 2018;9:3160.
https://doi.org/10.3389/fimmu.2018.03160 |
| 4 | Carreiro AL, Dhillon J, Gordon S, Higgins KA, Jacobs AG, McArthur BM, Redan BW, Rivera RL, Schmidt LR and Mattes RD: The Macronutrients, Appetite, and Energy Intake. Annu Rev Nutr 2016;36:73-103.
https://doi.org/10.1146/annurev-nutr-121415-112624 |
| 5 | Morris AL and Mohiuddin SS: Biochemistry, Nutrients: StatPearls. Treasure Island (FL) ineligible companies. Disclosure: Shamim Mohiuddin declares no relevant financial relationships with ineligible companies., 2025,
|
| 6 | Shenkin A: The key role of micronutrients. Clin Nutr 2006;25:1-13.
https://doi.org/10.1016/j.clnu.2005.11.006 |
| 7 | Stevens SL: Fat-soluble vitamins. Nursing Clinics 2021;56:33-45.
https://doi.org/10.1016/j.cnur.2020.10.003 |
| 8 | Gupta U and Gupta S: Sources and deficiency diseases of mineral nutrients in human health and nutrition: a review. Pedosphere 2014;24:13-38.
https://doi.org/10.1016/S1002-0160(13)60077-6 |
| 9 | Hamdan TA: The Multifaceted Roles of NK Cells in the Context of Murine Cytomegalovirus and Lymphocytic Choriomeningitis Virus Infections. Immune Netw 2024;24
https://doi.org/10.4110/in.2024.24.e29 |
| 10 | Hamdan TA, Ashraf F and Bhat H: Insights into Virus-Induced Immune Mediated Liver Pathology. Cell Physiol Biochem 2022;56:340-352.
https://doi.org/10.33594/000000554 |
| 11 | Duhan V, Hamdan TA, Xu HC, Shinde P, Bhat H, Li F, Al-Matary Y, Haussinger D, Bezgovsek J, Friedrich SK, Hardt C, Lang PA and Lang KS: NK cell-intrinsic FcepsilonRIgamma limits CD8+ T-cell expansion and thereby turns an acute into a chronic viral infection. PLoS Pathog 2019;15:e1007797.
https://doi.org/10.1371/journal.ppat.1007797 |
| 12 | Loftus RM, Assmann N, Kedia-Mehta N, O'Brien KL, Garcia A, Gillespie C, Hukelmann JL, Oefner PJ, Lamond AI, Gardiner CM, Dettmer K, Cantrell DA, Sinclair LV and Finlay DK: Amino acid-dependent cMyc expression is essential for NK cell metabolic and functional responses in mice. Nature Communications 2018;9:2341.
https://doi.org/10.1038/s41467-018-04719-2 |
| 13 | Arkan Yousif H, Al-Ani I, Hajleh MNA, Matalqah S, Dayyih WA and Al-Dujaili EA: Preparation and Evaluation of Complexed Ubiquinone (Coenzyme Q10) Antiaging Hyaluronic Acid-Vitamin C Serum for Skin Care. J Cosmet Dermatol 2025;24:e16706.
https://doi.org/10.1111/jocd.16706 |
| 14 | Nishikimi M, Fukuyama R, Minoshima S, Shimizu N and Yagi K: Cloning and chromosomal mapping of the human nonfunctional gene for L-gulono-gamma-lactone oxidase, the enzyme for L-ascorbic acid biosynthesis missing in man. J Biol Chem 1994;269:13685-8.
https://doi.org/10.1016/S0021-9258(17)36884-9 |
| 15 | Pawlowska E, Szczepanska J and Blasiak J: Pro- and Antioxidant Effects of Vitamin C in Cancer in correspondence to Its Dietary and Pharmacological Concentrations. Oxid Med Cell Longev 2019;2019:7286737.
https://doi.org/10.1155/2019/7286737 |
| 16 | Kim JE, Cho HS, Yang HS, Jung DJ, Hong SW, Hung CF, Lee WJ and Kim D: Depletion of ascorbic acid impairs NK cell activity against ovarian cancer in a mouse model. Immunobiology 2012;217:873-81.
https://doi.org/10.1016/j.imbio.2011.12.010 |
| 17 | Luchtel RA, Bhagat T, Pradhan K, Jacobs WR, Jr., Levine M, Verma A and Shenoy N: High-dose ascorbic acid synergizes with anti-PD1 in a lymphoma mouse model. Proc Natl Acad Sci U S A 2020;117:1666-1677.
https://doi.org/10.1073/pnas.1908158117 |
| 18 | Vojdani A and Ghoneum M: In vivo effect of ascorbic acid on enhancement of human natural killer cell activity. Nutrition Research 1993;13:753-764.
https://doi.org/10.1016/S0271-5317(05)80799-7 |
| 19 | Urbalejo-Ceniceros VI, Rocha-Gonzalez HI, Acosta-Maldonado BL, Valero-Saldana LM, Hernandez-Alcantara AE and Perez-Camargo DA: Effect of vitamin C on immune reconstitution after bone marrow transplantation. Int J Clin Pharmacol Ther 2022;60:384-391.
https://doi.org/10.5414/CP204182 |
| 20 | Huijskens MJ, Walczak M, Sarkar S, Atrafi F, Senden-Gijsbers BL, Tilanus MG, Bos GM, Wieten L and Germeraad WT: Ascorbic acid promotes proliferation of natural killer cell populations in culture systems applicable for natural killer cell therapy. Cytotherapy 2015;17:613-20.
https://doi.org/10.1016/j.jcyt.2015.01.004 |
| 21 | Kim H, Jang M, Kim Y, Choi J, Jeon J, Kim J, Hwang YI, Kang JS and Lee WJ: Red ginseng and vitamin C increase immune cell activity and decrease lung inflammation induced by influenza A virus/H1N1 infection. J Pharm Pharmacol 2016;68:406-20.
https://doi.org/10.1111/jphp.12529 |
| 22 | Peterson CT, Rodionov DA, Osterman AL and Peterson SN: B Vitamins and Their Role in Immune Regulation and Cancer. Nutrients 2020;12.
https://doi.org/10.3390/nu12113380 |
| 23 | Spinas E, Saggini A, Kritas SK, Cerulli G, Caraffa A, Antinolfi P, Pantalone A, Frydas A, Tei M, Speziali A, Saggini R, Pandolfi F and Conti P: Crosstalk between Vitamin B and Immunity. J Biol Regul Homeost Agents 2015;29:283-8.
|
| 24 | Tamura J, Kubota K, Murakami H, Sawamura M, Matsushima T, Tamura T, Saitoh T, Kurabayshi H and Naruse T: Immunomodulation by vitamin B12: augmentation of CD8+ T lymphocytes and natural killer (NK) cell activity in vitamin B12-deficient patients by methyl-B12 treatment. Clin Exp Immunol 1999;116:28-32.
https://doi.org/10.1046/j.1365-2249.1999.00870.x |
| 25 | Erkurt MA, Aydogdu I, Dikilitas M, Kuku I, Kaya E, Bayraktar N, Ozhan O, Ozkan I and Sonmez A: Effects of cyanocobalamin on immunity in patients with pernicious anemia. Med Princ Pract 2008;17:131-5.
https://doi.org/10.1159/000112967 |
| 26 | Mamcarz EK, Berg M, Peled T, Frei GM, Reger R, Tian X and Childs R: Nicotinamide (NAM) Increases Surface Expression of CD62L On in vitro Expanded NK Cells. Blood 2012;120:4355.
https://doi.org/10.1182/blood.V120.21.4355.4355 |
| 27 | Partearroyo T, Ubeda N, Montero A, Achon M and Varela-Moreiras G: Vitamin B(12) and folic acid imbalance modifies NK cytotoxicity, lymphocytes B and lymphoprolipheration in aged rats. Nutrients 2013;5:4836-48.
https://doi.org/10.3390/nu5124836 |
| 28 | Kim YI, Hayek M, Mason JB and Meydani SN: Severe folate deficiency impairs natural killer cell-mediated cytotoxicity in rats. J Nutr 2002;132:1361-7.
https://doi.org/10.1093/jn/132.6.1361 |
| 29 | Hirsch S, Miranda D, Fuentes C, Leiva L, Barrera G, Montoya M, Ronco AM, de la Maza MP and Bunout D: Effect of supraphysiological concentration of serum folate on natural killer cell activity in healthy subjects. e-SPEN Journal 2012;7:e125-e128.
https://doi.org/10.1016/j.clnme.2012.02.003 |
| 30 | Hirsch S, Miranda D, Muñoz E, Montoya M, Ronco AM, de la Maza MP and Bunout D: Natural killer cell cytotoxicity is not regulated by folic acid in vitro. Nutrition 2013;29:772-776.
https://doi.org/10.1016/j.nut.2012.10.006 |
| 31 | Ravaglia G, Forti P, Maioli F, Bastagli L, Facchini A, Mariani E, Savarino L, Sassi S, Cucinotta D and Lenaz G: Effect of micronutrient status on natural killer cell immune function in healthy free-living subjects aged >/=90 y. Am J Clin Nutr 2000;71:590-8.
https://doi.org/10.1093/ajcn/71.2.590 |
| 32 | Troen AM, Mitchell B, Sorensen B, Wener MH, Johnston A, Wood B, Selhub J, McTiernan A, Yasui Y, Oral E, Potter JD and Ulrich CM: Unmetabolized folic acid in plasma is associated with reduced natural killer cell cytotoxicity among postmenopausal women. J Nutr 2006;136:189-94.
https://doi.org/10.1093/jn/136.1.189 |
| 33 | Blomhoff R, Green MH, Berg T and Norum KR: Transport and storage of vitamin A. Science 1990;250:399-404.
https://doi.org/10.1126/science.2218545 |
| 34 | Colonna M: Interleukin-22-producing natural killer cells and lymphoid tissue inducer-like cells in mucosal immunity. Immunity 2009;31:15-23.
https://doi.org/10.1016/j.immuni.2009.06.008 |
| 35 | Tian Z, Chen Y and Gao B: Natural killer cells in liver disease. Hepatology 2013;57:1654-62.
https://doi.org/10.1002/hep.26115 |
| 36 | Jeong M, Cortopassi F, See JX, De La Torre C, Cerwenka A and Stojanovic A: Vitamin A-treated natural killer cells reduce interferon-gamma production and support regulatory T-cell differentiation. Eur J Immunol 2024;54:e2250342.
https://doi.org/10.1002/eji.202250342 |
| 37 | Zhao Z and Ross AC: Retinoic Acid Repletion Restores the Number of Leukocytes and Their Subsets and Stimulates Natural Cytotoxicity in Vitamin A-Deficient Rats. The Journal of Nutrition 1995;125:2064-2073.
https://doi.org/10.1093/jn/125.8.2064 |
| 38 | Bowman TA, Goonewardene IM, Pasatiempo AM, Ross AC and Taylor CE: Vitamin A deficiency decreases natural killer cell activity and interferon production in rats. J Nutr 1990;120:1264-73.
https://doi.org/10.1093/jn/120.10.1264 |
| 39 | Ross AC: Vitamin A deficiency and retinoid repletion regulate the antibody response to bacterial antigens and the maintenance of natural killer cells. Clin Immunol Immunopathol 1996;80:S63-72.
https://doi.org/10.1006/clin.1996.0143 |
| 40 | Fernandes-Carlos T, Riondel J, Glise D, Guiraud P and Favier A: Modulation of natural killer cell functional activity in athymic mice by beta-carotene, oestrone and their association. Anticancer Res 1997;17:2523-7.
|
| 41 | Carlos TF, Riondel J, Mathieu J, Guiraud P, Mestries JC and Favier A: Beta-carotene enhances natural killer cell activity in athymic mice. In vivo 1997;11:87-91.
|
| 42 | Jiang Y, Obuseh F, Ellis W, Piyathilake C and Jolly P: Association of vitamin A deficiency with decrease in TNF-alpha expressing CD3-CD56+ NK cells in Ghanaians. Nutr Res 2007;27:400-407.
https://doi.org/10.1016/j.nutres.2007.04.016 |
| 43 | Li A, He M, Wang H, Qiao B, Chen P, Gu H, Zhang M and He S: All-trans retinoic acid negatively regulates cytotoxic activities of nature killer cell line 92. Biochem Biophys Res Commun 2007;352:42-7.
https://doi.org/10.1016/j.bbrc.2006.10.132 |
| 44 | Lewis ED, Meydani SN and Wu D: Regulatory role of vitamin E in the immune system and inflammation. IUBMB Life 2019;71:487-494.
https://doi.org/10.1002/iub.1976 |
| 45 | Lee GY and Han SN: The Role of Vitamin E in Immunity. Nutrients 2018;10
https://doi.org/10.3390/nu10111614 |
| 46 | Traber MG: Vitamin E regulatory mechanisms. Annu Rev Nutr 2007;27:347-62.
https://doi.org/10.1146/annurev.nutr.27.061406.093819 |
| 47 | Cooper MA, Fehniger TA and Caligiuri MA: The biology of human natural killer-cell subsets. Trends Immunol 2001;22:633-40.
https://doi.org/10.1016/S1471-4906(01)02060-9 |
| 48 | Adachi N, Migita M, Ohta T, Higashi A and Matsuda I: Depressed natural killer cell activity due to decreased natural killer cell population in a vitamin E-deficient patient with Shwachman syndrome: reversible natural killer cell abnormality by alpha-tocopherol supplementation. Eur J Pediatr 1997;156:444-8.
https://doi.org/10.1007/s004310050634 |
| 49 | De la Fuente M, Hernanz A, Guayerbas N, Victor VM and Arnalich F: Vitamin E ingestion improves several immune functions in elderly men and women. Free Radic Res 2008;42:272-80.
https://doi.org/10.1080/10715760801898838 |
| 50 | Hanson MG, Ozenci V, Carlsten MC, Glimelius BL, Frodin JE, Masucci G, Malmberg KJ and Kiessling RV: A short-term dietary supplementation with high doses of vitamin E increases NK cell cytolytic activity in advanced colorectal cancer patients. Cancer Immunol Immunother 2007;56:973-84.
https://doi.org/10.1007/s00262-006-0261-4 |
| 51 | Silva ACO, Bonfim M, Fontes JLM, Dos-Santos WLC, Mengel J and Cardillo F: C57BL/6 Mice Pretreated With Alpha-Tocopherol Show a Better Outcome of Trypanosoma cruzi Infection With Less Tissue Inflammation and Fibrosis. Front Immunol 2022;13:833560.
https://doi.org/10.3389/fimmu.2022.833560 |
| 52 | Subramaniam S, Anandha Rao JS, Ramdas P, Ng MH, Kannan Kutty M, Selvaduray KR and Radhakrishnan AK: Reduced infiltration of regulatory T cells in tumours from mice fed daily with gamma-tocotrienol supplementation. Clin Exp Immunol 2021;206:161-172.
https://doi.org/10.1111/cei.13650 |
| 53 | Wang Y, Huang DS, Wood S and Watson RR: Modulation of immune function and cytokine production by various levels of vitamin E supplementation during murine AIDS. Immunopharmacology 1995;29:225-33.
https://doi.org/10.1016/0162-3109(95)00061-W |
| 54 | Abu Sa'aleek A, Alshishani A, Shaghlil L, Aljariri Alhesan JS and Al-ebini Y: Determination of vitamin D3 in pharmaceutical products using salting-out assisted liquid-liquid extraction coupled with reversed phase liquid chromatography. Microchemical Journal 2023;193:109077.
https://doi.org/10.1016/j.microc.2023.109077 |
| 55 | Perez-Lopez FR: Vitamin D: the secosteroid hormone and human reproduction. Gynecol Endocrinol 2007;23:13-24.
https://doi.org/10.1080/09513590601045629 |
| 56 | Raman M, Milestone AN, Walters JR, Hart AL and Ghosh S: Vitamin D and gastrointestinal diseases: inflammatory bowel disease and colorectal cancer. Therap Adv Gastroenterol 2011;4:49-62.
https://doi.org/10.1177/1756283X10377820 |
| 57 | Tripkovic L, Lambert H, Hart K, Smith CP, Bucca G, Penson S, Chope G, Hyppönen E, Berry J, Vieth R and Lanham-New S: Comparison of vitamin D2 and vitamin D3 supplementation in raising serum 25-hydroxyvitamin D status: a systematic review and meta-analysis123. The American Journal of Clinical Nutrition 2012;95:1357-1364.
https://doi.org/10.3945/ajcn.111.031070 |
| 58 | Cantorna MT and Mahon BD: Mounting evidence for vitamin D as an environmental factor affecting autoimmune disease prevalence. Exp Biol Med (Maywood) 2004;229:1136-42.
https://doi.org/10.1177/153537020422901108 |
| 59 | Khaled RA, Alhmoud JF, Issa RA, Khader HA, Mohammad BA, Alsayed AR, Khadra KA, Habash M, Aljaberi A, Hasoun LZ, Abu-Samak MS and Mosleh I: The variations of selected serum cytokines involved in cytokine Storm after Omega-3 daily supplements: A Randomized Clinical Trial in Jordanians with vitamin D deficiency. Pharmacy Practice 2024.
https://doi.org/10.18549/PharmPract.2024.2.2951 |
| 60 | Yu S and Cantorna MT: The vitamin D receptor is required for iNKT cell development. Proc Natl Acad Sci U S A 2008;105:5207-12.
https://doi.org/10.1073/pnas.0711558105 |
| 61 | Quesada JM, Serrano I, Borrego F, Martin A, Pena J and Solana R: Calcitriol effect on natural killer cells from hemodialyzed and normal subjects. Calcif Tissue Int 1995;56:113-7.
https://doi.org/10.1007/BF00296341 |
| 62 | Kitajima I, Maruyama I, Matsubara H, Osame M and Igata A: Immune dysfunction in hypophosphatemic vitamin D-resistant rickets: immunoregulatory reaction of 1 alpha(OH) vitamin D3. Clin Immunol Immunopathol 1989;53:24-31.
https://doi.org/10.1016/0090-1229(89)90097-4 |
| 63 | Vassiliou AG, Jahaj E, Pratikaki M, Keskinidou C, Detsika M, Grigoriou E, Psarra K, Orfanos SE, Tsirogianni A, Dimopoulou I and Kotanidou A: Vitamin D deficiency correlates with a reduced number of natural killer cells in intensive care unit (ICU) and non-ICU patients with COVID-19 pneumonia. Hellenic J Cardiol 2021;62:381-383.
https://doi.org/10.1016/j.hjc.2020.11.011 |
| 64 | Bychinin MV, Klypa TV, Mandel IA, Yusubalieva GM, Baklaushev VP, Kolyshkina NA and Troitsky AV: Effect of vitamin D3 supplementation on cellular immunity and inflammatory markers in COVID-19 patients admitted to the ICU. Sci Rep 2022;12:18604.
https://doi.org/10.1038/s41598-022-22045-y |
| 65 | Christofyllakis K, Neumann F, Bewarder M, Thurner L, Kaddu-Mulindwa D, Kos IA, Lesan V and Bittenbring JT: Vitamin D Enhances Immune Effector Pathways of NK Cells Thus Providing a Mechanistic Explanation for the Increased Effectiveness of Therapeutic Monoclonal Antibodies. Nutrients 2023;15.
https://doi.org/10.3390/nu15163498 |
| 66 | Berbudi A, Rahmadika N, Tjahjadi AI and Ruslami R: Type 2 Diabetes and its Impact on the Immune System. Curr Diabetes Rev 2020;16:442-449.
https://doi.org/10.2174/1573399815666191024085838 |
| 67 | Oh M, Jung S, Kim YA, Lee GY and Han SN: Dietary vitamin D(3) supplementation enhances splenic NK cell activity in healthy and diabetic male mice. Nutr Res 2024;127:144-155.
https://doi.org/10.1016/j.nutres.2024.06.004 |
| 68 | Weeres MA, Robien K, Ahn YO, Neulen ML, Bergerson R, Miller JS and Verneris MR: The effects of 1, 25-dihydroxyvitamin D3 on in vitro human NK cell development from hematopoietic stem cells. J Immunol 2014;193:3456-62.
https://doi.org/10.4049/jimmunol.1400698 |
| 69 | Ota K, Dambaeva S, Han AR, Beaman K, Gilman-Sachs A and Kwak-Kim J: Vitamin D deficiency may be a risk factor for recurrent pregnancy losses by increasing cellular immunity and autoimmunity. Hum Reprod 2014;29:208-19.
https://doi.org/10.1093/humrep/det424 |
| 70 | Ota K, Dambaeva S, Kim MW, Han AR, Fukui A, Gilman-Sachs A, Beaman K and Kwak-Kim J: 1, 25-Dihydroxy-vitamin D3 regulates NK-cell cytotoxicity, cytokine secretion, and degranulation in women with recurrent pregnancy losses. Eur J Immunol 2015;45:3188-99.
https://doi.org/10.1002/eji.201545541 |
| 71 | Gupta UC and Gupta SC: Sources and Deficiency Diseases of Mineral Nutrients in Human Health and Nutrition: A Review. Pedosphere 2014;24:13-38.
https://doi.org/10.1016/S1002-0160(13)60077-6 |
| 72 | Farag MA, Abib B, Qin Z, Ze X and Ali SE: Dietary macrominerals: Updated review of their role and orchestration in human nutrition throughout the life cycle with sex differences. Curr Res Food Sci 2023;6:100450.
https://doi.org/10.1016/j.crfs.2023.100450 |
| 73 | Weyh C, Kruger K, Peeling P and Castell L: The Role of Minerals in the Optimal Functioning of the Immune System. Nutrients 2022;14.
https://doi.org/10.3390/nu14030644 |
| 74 | Rajagopalan S and Long EO: Zinc bound to the killer cell-inhibitory receptor modulates the negative signal in human NK cells. J Immunol 1998;161:1299-305.
https://doi.org/10.4049/jimmunol.161.3.1299 |
| 75 | Rajagopalan S, Winter CC, Wagtmann N and Long EO: The Ig-related killer cell inhibitory receptor binds zinc and requires zinc for recognition of HLA-C on target cells. J Immunol 1995;155:4143-6.
https://doi.org/10.4049/jimmunol.155.9.4143 |
| 76 | Kumar S, Rajagopalan S, Sarkar P, Dorward DW, Peterson ME, Liao HS, Guillermier C, Steinhauser ML, Vogel SS and Long EO: Zinc-Induced Polymerization of Killer-Cell Ig-like Receptor into Filaments Promotes Its Inhibitory Function at Cytotoxic Immunological Synapses. Mol Cell 2016;62:21-33.
https://doi.org/10.1016/j.molcel.2016.03.009 |
| 77 | Wessels I, Maywald M and Rink L: Zinc as a Gatekeeper of Immune Function. Nutrients 2017;9.
https://doi.org/10.3390/nu9121286 |
| 78 | Maywald M and Rink L: Zinc supplementation dampens T helper 9 differentiation in allogeneic immune reactions in vitro. Unpublished work 2017.
https://doi.org/10.1016/j.jtemb.2018.02.004 |
| 79 | Muzzioli M, Stecconi R, Moresi R and Provinciali M: Zinc improves the development of human CD34+ cell progenitors towards NK cells and increases the expression of GATA-3 transcription factor in young and old ages. Biogerontology 2009;10:593-604.
https://doi.org/10.1007/s10522-008-9201-3 |
| 80 | Voli F, Valli E, Lerra L, Kimpton K, Saletta F, Giorgi FM, Mercatelli D, Rouaen JRC, Shen S, Murray JE, Ahmed-Cox A, Cirillo G, Mayoh C, Beavis PA, Haber M, Trapani JA, Kavallaris M and Vittorio O: Intratumoral Copper Modulates PD-L1 Expression and Influences Tumor Immune Evasion. Cancer Res 2020;80:4129-4144.
https://doi.org/10.1158/0008-5472.CAN-20-0471 |
| 81 | Donnelly RP, Loftus RM, Keating SE, Liou KT, Biron CA, Gardiner CM and Finlay DK: mTORC1-dependent metabolic reprogramming is a prerequisite for NK cell effector function. J Immunol 2014;193:4477-84.
https://doi.org/10.4049/jimmunol.1401558 |
| 82 | Littwitz-Salomon E, Moreira D, Frost JN, Choi C, Liou KT, Ahern DK, O'Shaughnessy S, Wagner B, Biron CA, Drakesmith H, Dittmer U and Finlay DK: Metabolic requirements of NK cells during the acute response against retroviral infection. Nat Commun 2021;12:5376.
https://doi.org/10.1038/s41467-021-25715-z |
| 83 | Hoffmann PR and Berry MJ: The influence of selenium on immune responses. Mol Nutr Food Res 2008;52:1273-80.
https://doi.org/10.1002/mnfr.200700330 |
| 84 | Wood SM, Beckham C, Yosioka A, Darban H and Watson RR: beta-Carotene and selenium supplementation enhances immune response in aged humans. Integr Med 2000;2:85-92.
https://doi.org/10.1016/S1096-2190(00)00009-3 |
| 85 | Lean MEJ: Principles of human nutrition. Medicine 2019;47:140-144.
https://doi.org/10.1016/j.mpmed.2018.12.014 |
| 86 | Mah AY, Rashidi A, Keppel MP, Saucier N, Moore EK, Alinger JB, Tripathy SK, Agarwal SK, Jeng EK, Wong HC, Miller JS, Fehniger TA, Mace EM, French AR and Cooper MA: Glycolytic requirement for NK cell cytotoxicity and cytomegalovirus control. JCI Insight 2017;2.
https://doi.org/10.1172/jci.insight.95128 |
| 87 | Pearce EL and Pearce EJ: Metabolic pathways in immune cell activation and quiescence. Immunity 2013;38:633-43.
https://doi.org/10.1016/j.immuni.2013.04.005 |
| 88 | Keppel MP, Saucier N, Mah AY, Vogel TP and Cooper MA: Activation-specific metabolic requirements for NK Cell IFN-gamma production. J Immunol 2015;194:1954-62.
https://doi.org/10.4049/jimmunol.1402099 |
| 89 | Marcais A, Cherfils-Vicini J, Viant C, Degouve S, Viel S, Fenis A, Rabilloud J, Mayol K, Tavares A, Bienvenu J, Gangloff YG, Gilson E, Vivier E and Walzer T: The metabolic checkpoint kinase mTOR is essential for IL-15 signaling during the development and activation of NK cells. Nat Immunol 2014;15:749-757.
https://doi.org/10.1038/ni.2936 |
| 90 | Assmann N, O'Brien KL, Donnelly RP, Dyck L, Zaiatz-Bittencourt V, Loftus RM, Heinrich P, Oefner PJ, Lynch L, Gardiner CM, Dettmer K and Finlay DK: Srebp-controlled glucose metabolism is essential for NK cell functional responses. Nat Immunol 2017;18:1197-1206.
https://doi.org/10.1038/ni.3838 |
| 91 | Picard LK, Littwitz-Salomon E, Waldmann H and Watzl C: Inhibition of Glucose Uptake Blocks Proliferation but Not Cytotoxic Activity of NK Cells. Cells 2022;11.
https://doi.org/10.3390/cells11213489 |
| 92 | Kim JH, Park K, Lee SB, Kang S, Park JS, Ahn CW and Nam JS: Relationship between natural killer cell activity and glucose control in patients with type 2 diabetes and prediabetes. J Diabetes Investig 2019;10:1223-1228.
https://doi.org/10.1111/jdi.13002 |
| 93 | Berrou J, Fougeray S, Venot M, Chardiny V, Gautier JF, Dulphy N, Toubert A and Peraldi MN: Natural killer cell function, an important target for infection and tumor protection, is impaired in type 2 diabetes. PLoS One 2013;8:e62418.
https://doi.org/10.1371/journal.pone.0062418 |
| 94 | Zhang Y, Wang H, Lou X, Qu X, Gao L, Liu X, Li M, Guo H and Jiang Y: Decreased percentage of NKG2D+NK cells in patients with incident onset of Type 1 Diabetes. Clin Exp Pharmacol Physiol 2017;44:180-190.
https://doi.org/10.1111/1440-1681.12699 |
| 95 | Li P, Yin YL, Li D, Kim SW and Wu G: Amino acids and immune function. Br J Nutr 2007;98:237-52.
https://doi.org/10.1017/S000711450769936X |
| 96 | Ruth MR and Field CJ: The immune modifying effects of amino acids on gut-associated lymphoid tissue. J Anim Sci Biotechnol 2013;4:27.
https://doi.org/10.1186/2049-1891-4-27 |
| 97 | Field CJ, Johnson IR and Schley PD: Nutrients and their role in host resistance to infection. J Leukoc Biol 2002;71:16-32.
https://doi.org/10.1189/jlb.71.1.16 |
| 98 | Vicario M, Amat C, Rivero M, Moreto M and Pelegri C: Dietary glutamine affects mucosal functions in rats with mild DSS-induced colitis. J Nutr 2007;137:1931-7.
https://doi.org/10.1093/jn/137.8.1931 |
| 99 | Song QH, Xu RM, Zhang QH, Shen GQ, Ma M, Zhao XP, Guo YH and Wang Y: Glutamine supplementation and immune function during heavy load training. Int J Clin Pharmacol Ther 2015;53:372-6.
https://doi.org/10.5414/CP202227 |
| 100 | Krzywkowski K, Petersen EW, Ostrowski K, Kristensen JH, Boza J and Pedersen BK: Effect of glutamine supplementation on exercise-induced changes in lymphocyte function. Am J Physiol Cell Physiol 2001;281:C1259-65.
https://doi.org/10.1152/ajpcell.2001.281.4.C1259 |
| 101 | Bhat ZF, Kumar S and Bhat HF: Bioactive peptides of animal origin: a review. Journal of Food Science and Technology 2015;52:5377-5392.
https://doi.org/10.1007/s13197-015-1731-5 |
| 102 | Chalamaiah M, Yu W and Wu J: Immunomodulatory and anticancer protein hydrolysates (peptides) from food proteins: A review. Food Chem 2018;245:205-222.
https://doi.org/10.1016/j.foodchem.2017.10.087 |
| 103 | Cai B, Pan J, Wu Y, Wan P and Sun H: Immune functional impacts of oyster peptide-based enteral nutrition formula (OPENF) on mice: a pilot study. Chinese Journal of Oceanology and Limnology 2013;31:813-820.
https://doi.org/10.1007/s00343-013-2311-z |
| 104 | Wang Y-K, He H-L, Wang G-F, Wu H, Zhou B-C, Chen X-L and Zhang Y-Z: Oyster (Crassostrea gigas) Hydrolysates Produced on a Plant Scale Have Antitumor Activity and Immunostimulating Effects in BALB/c Mice. Marine Drugs. DOI: 10.3390/md8020255.
https://doi.org/10.3390/md8020255 |
| 105 | Chalamaiah M, Hemalatha R, Jyothirmayi T, Diwan PV, Bhaskarachary K, Vajreswari A, Ramesh Kumar R and Dinesh Kumar B: Chemical composition and immunomodulatory effects of enzymatic protein hydrolysates from common carp (Cyprinus carpio) egg. Nutrition 2015;31:388-398.
https://doi.org/10.1016/j.nut.2014.08.006 |
| 106 | Chalamaiah M, Hemalatha R, Jyothirmayi T, Diwan PV, Uday Kumar P, Nimgulkar C and Dinesh Kumar B: Immunomodulatory effects of protein hydrolysates from rohu (Labeo rohita) egg (roe) in BALB/c mice. Food Research International 2014;62:1054-1061.
https://doi.org/10.1016/j.foodres.2014.05.050 |
| 107 | Yaqoob P and Calder PC: Fatty acids and immune function: new insights into mechanisms. Br J Nutr 2007;98 Suppl 1:S41-5.
https://doi.org/10.1017/S0007114507832995 |
| 108 | Sheppard S, Srpan K, Lin W, Lee M, Delconte RB, Owyong M, Carmeliet P, Davis DM, Xavier JB, Hsu KC and Sun JC: Fatty acid oxidation fuels natural killer cell responses against infection and cancer. Proc Natl Acad Sci U S A 2024;121:e2319254121.
https://doi.org/10.1073/pnas.2319254121 |
| 109 | Manzo T, Prentice BM, Anderson KG, Raman A, Schalck A, Codreanu GS, Nava Lauson CB, Tiberti S, Raimondi A, Jones MA, Reyzer M, Bates BM, Spraggins JM, Patterson NH, McLean JA, Rai K, Tacchetti C, Tucci S, Wargo JA, Rodighiero S, Clise-Dwyer K, Sherrod SD, Kim M, Navin NE, Caprioli RM, Greenberg PD, Draetta G and Nezi L: Accumulation of long-chain fatty acids in the tumor microenvironment drives dysfunction in intrapancreatic CD8+ T cells. J Exp Med 2020;217.
https://doi.org/10.1084/jem.20191920 |
| 110 | Xu S, Chaudhary O, Rodriguez-Morales P, Sun X, Chen D, Zappasodi R, Xu Z, Pinto AFM, Williams A, Schulze I, Farsakoglu Y, Varanasi SK, Low JS, Tang W, Wang H, McDonald B, Tripple V, Downes M, Evans RM, Abumrad NA, Merghoub T, Wolchok JD, Shokhirev MN, Ho PC, Witztum JL, Emu B, Cui G and Kaech SM: Uptake of oxidized lipids by the scavenger receptor CD36 promotes lipid peroxidation and dysfunction in CD8(+) T cells in tumors. Immunity 2021;54:1561-1577 e7.
https://doi.org/10.1016/j.immuni.2021.05.003 |
| 111 | Schoors S, Bruning U, Missiaen R, Queiroz KC, Borgers G, Elia I, Zecchin A, Cantelmo AR, Christen S, Goveia J, Heggermont W, Godde L, Vinckier S, Van Veldhoven PP, Eelen G, Schoonjans L, Gerhardt H, Dewerchin M, Baes M, De Bock K, Ghesquiere B, Lunt SY, Fendt SM and Carmeliet P: Fatty acid carbon is essential for dNTP synthesis in endothelial cells. Nature 2015;520:192-197.
https://doi.org/10.1038/nature14362 |
| 112 | Delconte RB, Owyong M, Santosa EK, Srpan K, Sheppard S, McGuire TJ, Abbasi A, Diaz-Salazar C, Chun J, Rogatsky I, Hsu KC, Jordan S, Merad M and Sun JC: Fasting reshapes tissue-specific niches to improve NK cell-mediated anti-tumor immunity. Immunity 2024;57:1923-1938.e7.
https://doi.org/10.1016/j.immuni.2024.05.021 |
| 113 | Schimmer S, Mittermuller D, Werner T, Gors PE, Meckelmann SW, Finlay DK, Dittmer U and Littwitz-Salomon E: Fatty acids are crucial to fuel NK cells upon acute retrovirus infection. Front Immunol 2023;14:1296355.
https://doi.org/10.3389/fimmu.2023.1296355 |
| 114 | Xie Z, Fu Y, Tan WS and Cai H: Fatty acids promote the expansion of NK-92 cells in vitro by improving energy metabolism. Appl Microbiol Biotechnol 2021;105:4285-4295.
https://doi.org/10.1007/s00253-021-11313-y |
| 115 | Kobayashi T, Lam PY, Jiang H, Bednarska K, Gloury R, Murigneux V, Tay J, Jacquelot N, Li R, Tuong ZK, Leggatt GR, Gandhi MK, Hill MM, Belz GT, Ngo S, Kallies A and Mattarollo SR: Increased lipid metabolism impairs NK cell function and mediates adaptation to the lymphoma environment. Blood 2020;136:3004-3017.
https://doi.org/10.1182/blood.2020005602 |
| 116 | Michelet X, Dyck L, Hogan A, Loftus RM, Duquette D, Wei K, Beyaz S, Tavakkoli A, Foley C, Donnelly R, O'Farrelly C, Raverdeau M, Vernon A, Pettee W, O'Shea D, Nikolajczyk BS, Mills KHG, Brenner MB, Finlay D and Lynch L: Metabolic reprogramming of natural killer cells in obesity limits antitumor responses. Nat Immunol 2018;19:1330-1340.
https://doi.org/10.1038/s41590-018-0251-7 |
| 117 | Bahr I, Goritz V, Doberstein H, Hiller GG, Rosenstock P, Jahn J, Portner O, Berreis T, Mueller T, Spielmann J and Kielstein H: Diet-Induced Obesity Is Associated with an Impaired NK Cell Function and an Increased Colon Cancer Incidence. J Nutr Metab 2017;2017:4297025.
https://doi.org/10.1155/2017/4297025 |
| 118 | Spielmann J, Hanke J, Knauf D, Ben-Eliyahu S, Jacobs R, Stangl GI, Bahr I and Kielstein H: Significantly enhanced lung metastasis and reduced organ NK cell functions in diet-induced obese rats. BMC Obes 2017;4:24.
https://doi.org/10.1186/s40608-017-0161-5 |
| 119 | Spielmann J, Mattheis L, Jung JS, Rausse H, Glass M, Bahr I, Quandt D, Oswald J and Kielstein H: Effects of obesity on NK cells in a mouse model of postmenopausal breast cancer. Sci Rep 2020;10:20606.
https://doi.org/10.1038/s41598-020-76906-5 |
| 120 | Spielmann J, Naujoks W, Emde M, Allweyer M, Kielstein H, Quandt D and Bahr I: High-Fat Diet and Feeding Regime Impairs Number, Phenotype, and Cytotoxicity of Natural Killer Cells in C57BL/6 Mice. Front Nutr 2020;7:585693.
https://doi.org/10.3389/fnut.2020.585693 |
| 121 | Cacalano NA: Regulation of Natural Killer Cell Function by STAT3. Front Immunol 2016;7:128.
https://doi.org/10.3389/fimmu.2016.00128 |
| 122 | Agura T, Shin S, Jo H, Jeong S, Ahn H, Pang SY, Lee J, Park JH, Kim Y and Kang JS: Aptamin C enhances anti-cancer activity NK cells through the activation of STAT3: a comparative study with vitamin C. Anat Cell Biol 2024;57:408-418.
https://doi.org/10.5115/acb.24.120 |
| 123 | Heuser G and Vojdani A: Enhancement of natural killer cell activity and T and B cell function by buffered vitamin C in patients exposed to toxic chemicals: the role of protein kinase-C. Immunopharmacol Immunotoxicol 1997;19:291-312.
https://doi.org/10.3109/08923979709046977 |
| 124 | Sadeghi K, Wessner B, Laggner U, Ploder M, Tamandl D, Friedl J, Zugel U, Steinmeyer A, Pollak A, Roth E, Boltz-Nitulescu G and Spittler A: Vitamin D3 down-regulates monocyte TLR expression and triggers hyporesponsiveness to pathogen-associated molecular patterns. Eur J Immunol 2006;36:361-70.
https://doi.org/10.1002/eji.200425995 |
| 125 | Mian MF, Lauzon NM, Andrews DW, Lichty BD and Ashkar AA: FimH can directly activate human and murine natural killer cells via TLR4. Mol Ther 2010;18:1379-88.
https://doi.org/10.1038/mt.2010.75 |
| 126 | Loftus RM, Assmann N, Kedia-Mehta N, O'Brien KL, Garcia A, Gillespie C, Hukelmann JL, Oefner PJ, Lamond AI, Gardiner CM, Dettmer K, Cantrell DA, Sinclair LV and Finlay DK: Amino acid-dependent cMyc expression is essential for NK cell metabolic and functional responses in mice. Nat Commun 2018;9:2341.
https://doi.org/10.1038/s41467-018-04719-2 |
| 127 | Rodacki M, Svoren B, Butty V, Besse W, Laffel L, Benoist C and Mathis D: Altered natural killer cells in type 1 diabetic patients. Diabetes 2007;56:177-85.
https://doi.org/10.2337/db06-0493 |
| 128 | Conde-Knape K: Heparan sulfate proteoglycans in experimental models of diabetes: a role for perlecan in diabetes complications. Diabetes Metab Res Rev 2001;17:412-21.
https://doi.org/10.1002/dmrr.236 |
| 129 | Bloushtain N, Qimron U, Bar-Ilan A, Hershkovitz O, Gazit R, Fima E, Korc M, Vlodavsky I, Bovin NV and Porgador A: Membrane-associated heparan sulfate proteoglycans are involved in the recognition of cellular targets by NKp30 and NKp46. J Immunol 2004;173:2392-401.
https://doi.org/10.4049/jimmunol.173.4.2392 |
| 130 | Hirsch S, Miranda D, Munoz E, Montoya M, Ronco AM, de la Maza MP and Bunout D: Natural killer cell cytotoxicity is not regulated by folic acid in vitro. Nutrition 2013;29:772-6.
https://doi.org/10.1016/j.nut.2012.10.006 |
| 131 | Amento EP, Bhalla AK, Kurnick JT, Kradin RL, Clemens TL, Holick SA, Holick MF and Krane SM: 1 alpha,25-dihydroxyvitamin D3 induces maturation of the human monocyte cell line U937, and, in association with a factor from human T lymphocytes, augments production of the monokine, mononuclear cell factor. J Clin Invest 1984;73:731-9.
https://doi.org/10.1172/JCI111266 |
| 132 | Mahon BD, Wittke A, Weaver V and Cantorna MT: The targets of vitamin D depend on the differentiation and activation status of CD4 positive T cells. J Cell Biochem 2003;89:922-32.
https://doi.org/10.1002/jcb.10580 |
| 133 | He C, Wang D, Shukla SK, Hu T, Thakur R, Fu X, King RJ, Kollala SS, Attri KS, Murthy D, Chaika NV, Fujii Y, Gonzalez D, Pacheco CG, Qiu Y, Singh PK, Locasale JW and Mehla K: Vitamin B6 Competition in the Tumor Microenvironment Hampers Antitumor Functions of NK Cells. Cancer Discov 2024;14:176-193.
https://doi.org/10.1158/2159-8290.CD-23-0334 |
| 134 | Zhang K and Kaufman RJ: From endoplasmic-reticulum stress to the inflammatory response. Nature 2008;454:455-62.
https://doi.org/10.1038/nature07203 |
| 135 | Peraldi MN, Berrou J, Dulphy N, Seidowsky A, Haas P, Boissel N, Metivier F, Randoux C, Kossari N, Guerin A, Geffroy S, Delavaud G, Marin-Esteban V, Glotz D, Charron D and Toubert A: Oxidative stress mediates a reduced expression of the activating receptor NKG2D in NK cells from end-stage renal disease patients. J Immunol 2009;182:1696-705.
https://doi.org/10.4049/jimmunol.182.3.1696 |
| 136 | Huntington ND, Legrand N, Alves NL, Jaron B, Weijer K, Plet A, Corcuff E, Mortier E, Jacques Y, Spits H and Di Santo JP: IL-15 trans-presentation promotes human NK cell development and differentiation in vivo. J Exp Med 2009;206:25-34.
https://doi.org/10.1084/jem.20082013 |
| 137 | Genuardi JA, Eskew ML, Zeligs BJ and Bellanti JA: The Effects of Vitamin E and Selenium on The Nitric Oxide Production of Macrophages. Pediatric Research 1999;45:771-771.
https://doi.org/10.1203/00006450-199905010-00201 |
| 138 | Furuke K, Burd PR, Horvath-Arcidiacono JA, Hori K, Mostowski H and Bloom ET: Human NK cells express endothelial nitric oxide synthase, and nitric oxide protects them from activation-induced cell death by regulating expression of TNF-alpha. J Immunol 1999;163:1473-80.
https://doi.org/10.4049/jimmunol.163.3.1473 |
| 139 | Cifone MG, D'Alo S, Parroni R, Millimaggi D, Biordi L, Martinotti S and Santoni A: Interleukin-2-activated rat natural killer cells express inducible nitric oxide synthase that contributes to cytotoxic function and interferon-gamma production. Blood 1999;93:3876-84.
https://doi.org/10.1182/blood.V93.11.3876.411k25_3876_3884 |
| 140 | Stiff A, Trikha P, Mundy-Bosse B, McMichael E, Mace TA, Benner B, Kendra K, Campbell A, Gautam S, Abood D, Landi I, Hsu V, Duggan M, Wesolowski R, Old M, Howard JH, Yu L, Stasik N, Olencki T, Muthusamy N, Tridandapani S, Byrd JC, Caligiuri M and Carson WE: Nitric Oxide Production by Myeloid-Derived Suppressor Cells Plays a Role in Impairing Fc Receptor-Mediated Natural Killer Cell Function. Clin Cancer Res 2018;24:1891-1904.
https://doi.org/10.1158/1078-0432.CCR-17-0691 |
| 141 | Young JD, Leong LG, Liu CC, Damiano A and Cohn ZA: Extracellular release of lymphocyte cytolytic pore-forming protein (perforin) after ionophore stimulation. Proc Natl Acad Sci U S A 1986;83:5668-72.
https://doi.org/10.1073/pnas.83.15.5668 |