Ligament Cell Biology: Effect of Mechanical Loading
bInterdisciplinary Scientific Group of Sports Medicine, Department of Sports Medicine, Medical University of Lublin, Lublin, Poland,
cInstitute of Physiotherapy and Health Sciences, The Jerzy Kukuczka Academy of Physical Education, Katowice, Poland,
dFunctional Diagnostics Laboratory, Sport-Klinika, Scanmed Sport, Żory, Poland,
eSports Research Center Nicolaus Copernicus University, Toruń, Poland
Keywords
Abstract
Ligaments are biomechanically specialized connective tissues that maintain joint stability and guide motion under complex loading conditions. At the cellular and molecular levels, ligament homeostasis is governed by fibroblast-like cells (ligamentocytes) embedded in an intricately organized ECM composed predominantly of type I collagen, with contributions from type III collagen, elastin, proteoglycans, and glycoproteins. These cells continuously sense and respond to mechanical stimuli—tension, compression, and shear—through mechanotransduction pathways involving integrins, focal adhesions, cytoskeletal remodeling, and mechanosensitive ion channels. Downstream signaling cascades, including MAPKs and PI3K/AKT, integrate biomechanical cues with growth factor and cytokine signaling to fine-tune gene expression, collagen fibrillogenesis, and ECM turnover. Distinct from tendons, ligaments must adapt to multidirectional loads, resulting in unique ECM compositions and cellular phenotypes. Appropriate mechanical loading maintains collagen alignment, promotes ECM integrity, and stabilizes the ligament cell phenotype. By contrast, insufficient or excessive load alters the molecular balance, triggering catabolic processes, inflammation, and disorganized ECM assembly. This delicate equilibrium also underlies the ligamentization observed in ACL graft remodeling, where controlled mechanical environments and molecular interventions accelerate the acquisition of ligamentous properties. Emerging insights into transcriptional and epigenetic regulation, growth factor-mediated cues, and cytokine-driven responses offer avenues to engineer ligament-like tissues and optimize recovery strategies. By leveraging molecular knowledge of cell–matrix interactions, growth factor profiles, and genetic/epigenetic modulators, clinicians and researchers can design tailored loading protocols, biomimetic scaffolds, and regenerative therapies. These approaches aim to restore ligament functionality, enhance graft integration, and prevent degenerative changes, ultimately improving patient outcomes in ligament injury repair and reconstruction.
Introduction
Ligaments are critical connective tissues that link bones and guide joint motion, withstanding complex, multidirectional mechanical loads that ensure joint stability and proper arthrokinematics [1–3]. At the cellular and molecular levels, ligament biology is distinguished by a specialized population of fibroblast-like cells, often termed ligament fibroblasts or ligamentocytes, that reside within a highly organized extracellular matrix (ECM) [4–7]. The ECM of ligaments is composed predominantly of type I collagen, which provides tensile strength, but also includes type III collagen, elastin, and an array of proteoglycans and glycoproteins that contribute to viscoelastic behavior and tissue compliance [8–12]. This diverse molecular environment allows ligaments to fulfill their demanding mechanical functions under various loading regimes [13–15].
Within the ECM milieu, ligament fibroblasts actively sense and respond to mechanical cues through mechanotransduction pathways [16–19]. Mechanical loading—whether tensile stretch during extension, shear forces during rotational movements, or transient compressive loads during joint impact—modulates a cascade of intracellular signals that ultimately influence gene expression, protein synthesis, and ECM remodeling [20–23]. At the molecular level, integrins—transmembrane receptors that anchor cells to collagen fibrils—serve as primary sensors of mechanical forces and transmit signals to focal adhesion complexes [24–26]. These complexes include cytoskeletal adaptor proteins, such as vinculin and talin, which link integrin receptors to the actin cytoskeleton [27–29]. Changes in tension and matrix stiffness alter integrin clustering and conformation, triggering the activation of focal adhesion kinase (FAK) and downstream signaling molecules including mitogen-activated protein kinases (MAPKs) and PI3K/AKT pathways [30–33]. In parallel, ligament fibroblasts detect mechanical perturbations via mechanosensitive ion channels, stretch-activated receptors, and the deformation of primary cilia, all of which converge upon intracellular signal transduction networks [34–37]. These signals dictate the transcriptional regulation of structural proteins (e.g., collagen I, collagen III), proteoglycans (decorin, biglycan), and matrix-remodeling enzymes (e.g., matrix metalloproteinases), as well as growth factors and cytokines that modulate cellular proliferation, differentiation, and ECM turnover [38–42]. Additionally, dynamic changes in ECM composition influence ligand binding and growth factor sequestration, further refining the molecular interplay between cells and their surrounding matrix [43–45]. This intricate relationship ensures that ligament fibroblasts continuously adapt their molecular and cellular states in response to the mechanical environment [46–48]. When subjected to appropriate levels of physiologic load, these cells maintain homeostasis—preserving proper collagen fibril alignment, ECM integrity, and tissue mechanical properties [49–51]. Conversely, deviations in loading conditions, such as excessive strain or prolonged unloading, can elicit maladaptive cellular responses, leading to ECM disorganization, altered collagen fiber composition, increased proteolytic enzyme activity, and a shift towards a more fibrocartilaginous phenotype [52–55]. Such maladaptive changes underlie ligament degeneration, impaired healing, and compromised mechanical function following injury [56–58].
A nuanced understanding of ligament cell mechanobiology is, therefore, pivotal for informing clinical strategies aimed at restoring normal tissue function after injury, enhancing rehabilitation protocols, and advancing tissue engineering approaches that mimic the native mechanical microenvironment [59–61]. By manipulating molecular signaling pathways, fine-tuning mechanical stimulation parameters, and leveraging emerging biotechnologies—such as growth factor delivery, gene editing, or biomimetic scaffolds—it may be possible to direct ligament fibroblasts toward desirable phenotypes [62–64]. Ultimately, improving our knowledge of the molecular and cellular responses to mechanical loading will contribute significantly to the prevention of ligament injuries, the optimization of post-injury treatement, and the successful engineering of functional ligamentous tissues [65–67].
Molecular and Cellular Differences Between Ligaments and Tendons
While ligaments and tendons share fundamental structural and hierarchical organization—both are dense, collagen-rich connective tissues connecting musculoskeletal components—their molecular profiles and cellular phenotypes diverge to accommodate distinct biomechanical roles (Table 1). Tendons typically transfer unidirectional tensile loads from muscle to bone, resulting in a highly aligned, parallel array of type I collagen fibers with minimal elastin and relatively sparse proteoglycan content [68–71]. This predominance of type I collagen, coupled with low levels of glycosaminoglycans (GAGs), reflects an adaptation to resist tensile loading along a single predominant axis. Ligaments, in contrast, withstand more complex, multidirectional mechanical environments associated with joint stabilization. As a result, ligament ECM composition frequently includes a higher proportion of type III collagen interspersed among the predominant type I collagen fibrils, as well as greater quantities of elastin and small leucine-rich proteoglycans (SLRPs) such as decorin, biglycan, fibromodulin, and lumican [72–74]. These molecules modulate collagen fibrillogenesis, influence fibril diameter distribution, and contribute to the viscoelastic properties needed to endure variable loading states. The presence of elastin and a richer array of proteoglycans imparts ligaments with greater compliance and an enhanced ability to absorb and dissipate complex stresses, improving joint stability while allowing controlled mobility. At the cellular level, ligament fibroblasts (ligamentocytes) differ from tendon fibroblasts (tenocytes) in their gene expression patterns and responsiveness to mechanical signals [75–77]. Ligament cells express a distinct repertoire of integrins, including integrin subunits that may preferentially bind to ECM components enriched in ligaments, influencing downstream signaling pathways that guide cytoskeletal organization and ECM remodeling. For instance, certain α- and β-integrin subunits may be upregulated in ligament fibroblasts to facilitate adhesion to collagen III- and proteoglycan-rich matrices, while tendon cells rely more heavily on integrins tuned to linear collagen I networks. Additionally, ligament fibroblasts produce higher levels of elastin and glycoproteins that enhance tissue compliance and enable load distribution across multiple axes. Their secretory profile often includes distinct growth factors (e.g., TGF-β isoforms, FGF, and CTGF) and cytokines that reflect the need for dynamic remodeling under complex loading conditions. These signaling molecules not only regulate collagen synthesis and degradation but also modulate matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs), balancing anabolic and catabolic activities to maintain ligament homeostasis.
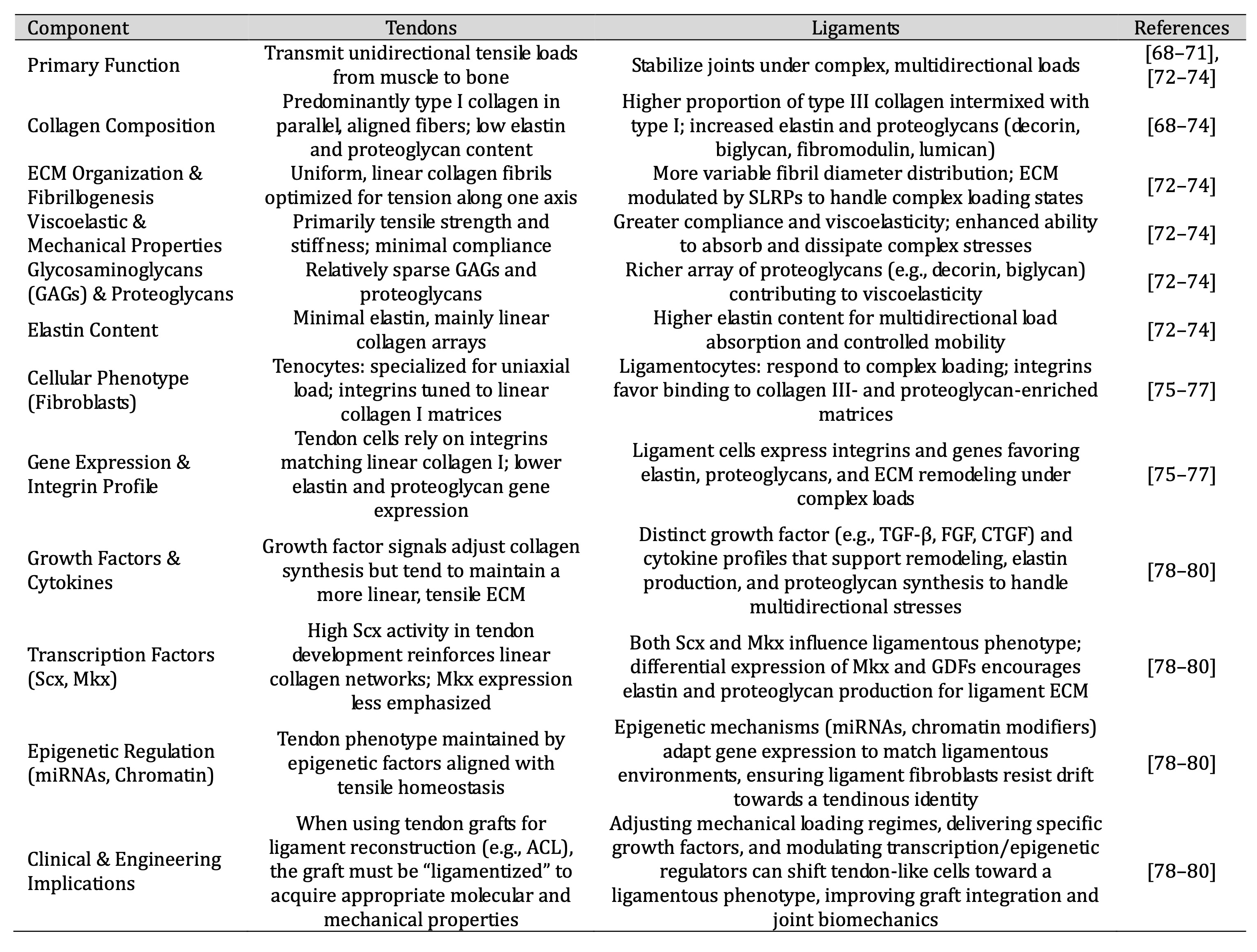
Table 1: This table highlights how tendons and ligaments, despite both being collagen-rich connective tissues, differ in ECM composition, mechanical properties, cellular phenotypes, integrin expression, and transcriptional/epigenetic regulation. These distinctions form the basis for targeted regenerative medicine approaches that manipulate molecular and cellular factors to steer tendon grafts and engineered constructs toward a ligamentous phenotype, ultimately improving clinical outcomes in ligament repair and reconstruction
Key transcription factors such as scleraxis (Scx) and Mohawk (Mkx) are central to both tendon and ligament development, but subtle variations in their expression, as well as their interaction with cofactors like EGR1, GDFs, and other regulatory proteins, can steer cells toward a more ligamentous or more tendinous phenotype [78–80]. For example, differential expression levels of Mkx and GDFs might favor the production of elastin and proteoglycans characteristic of ligament ECM, while high Scx activity in concert with different co-regulators might reinforce a strictly linear collagen network more akin to tendon tissue.
Epigenetic regulators, including microRNAs and chromatin-modifying enzymes, may also differ between ligament and tendon fibroblasts, fine-tuning gene expression to match the unique mechanical and biochemical environments of each tissue. These epigenetic factors can govern the responsiveness to growth factors, integrin-mediated signaling, and cytoskeletal stress, ensuring that the cells retain their ligamentous identity and avoid drifting toward a tendinous phenotype—or vice versa.
Understanding these molecular and cellular distinctions is crucial for regenerative medicine and tissue engineering strategies. When using tendon grafts to reconstruct ruptured ligaments (e.g., the anterior cruciate ligament), it is essential that the transplanted tissue acquires ligament-like molecular attributes. By manipulating mechanical loading parameters, delivering specific growth factors, or modulating transcriptional and epigenetic regulators, it may be possible to guide tendon cells toward a ligament-like phenotype. This level of molecular control can improve graft integration and ultimately enhance clinical outcomes after ligament reconstruction procedures.
Mechanotransduction Pathways in Ligament Cells
Mechanotransduction—the process by which ligament fibroblasts convert mechanical stimuli into biochemical and genetic responses—underpins the dynamic remodeling and maintenance of connective tissues subjected to complex, multiaxial loads [81–83] (Fig. 1). In ligaments, this involves a finely orchestrated network comprising integrin receptors, mechanosensitive ion channels, G protein-coupled receptors (GPCRs), primary cilia, cytoskeletal filaments, and multiple downstream molecular effectors. Collectively, these sensors and signaling hubs regulate the synthesis, organization, and turnover of the extracellular ECM. Additionally, specific genetic variants such as R2482H can modify key components of calcium-dependent signaling via calcineurin—sometimes referred to as “calcineurin 3”—and transcription factors of the NFATC family, imparting unique susceptibility or adaptive responses in ligament tissues. Integrins, which exist as transmembrane α/β heterodimers, physically couple ligament fibroblasts to collagen fibrils, proteoglycans, and glycoproteins within the ECM [84–86]. Mechanical strain alters the conformation and clustering of integrin subunits (e.g., α5β1, αvβ3), triggering the recruitment of focal adhesion proteins such as talin, vinculin, paxillin, and α-actinin to nascent adhesion complexes [87–89]. Upon integrin activation, FAK undergoes phosphorylation, creating scaffolding sites for Src family kinases and other intermediates that converge on MAPK (ERK, JNK, p38) and PI3K/AKT pathways [30–32] [90–92]. ERK commonly mediates proliferation and collagen-related gene expression, while JNK and p38 act as stress-activated kinases, reorganizing the actin cytoskeleton and modulating MMPs. Meanwhile, PI3K/AKT coordinates cell survival and metabolic adaptation, allowing ligament fibroblasts to calibrate collagen deposition, proteoglycan assembly, and fibril alignment according to the mechanical demands imposed on the tissue.
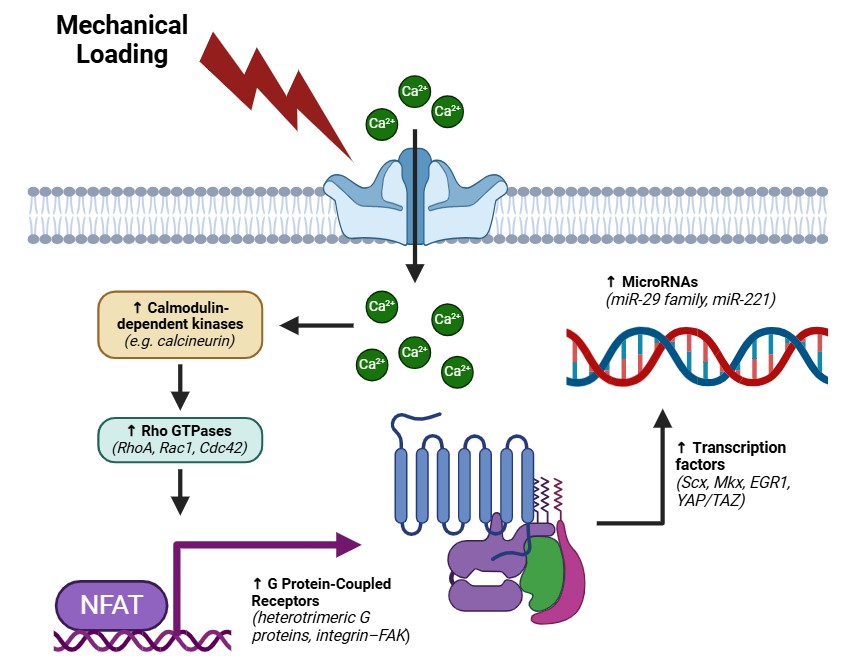
Fig. 1: Graphical representation of ligament mechanosensing. This figure illustrates the principal mechanosensitive pathways that operate within ligament fibroblasts following mechanical loading. At the cell–ECM interface, mechanical strain on integrins (e.g., α5β1, αvβ3) triggers clustering of focal adhesion proteins—such as talin, vinculin, paxillin, and α-actinin—thereby activating and phosphorylating focal adhesion kinase (FAK). In turn, FAK provides a platform for Src kinases and adaptors that merge into two major downstream pathways: the MAPK cascade (ERK, JNK, p38) and the PI3K/AKT axis. ERK drives cell proliferation and collagen gene expression, whereas JNK and p38 mediate cytoskeletal remodeling and MMP regulation. The PI3K/AKT pathway orchestrates cell survival signals and modulates collagen deposition, proteoglycan assembly, and fibril alignment.Meanwhile, mechanosensitive ion channels (e.g., TRPV4 and PIEZO1) detect membrane deformation and facilitate rapid Ca²⁺ influx, activating calmodulin-dependent kinases or phosphatases (e.g., calcineurin). Elevated intracellular Ca²⁺ also influences Rho GTPases (RhoA, Rac1, Cdc42), promoting changes in actin filament organization and focal adhesion architecture. These modifications alter nuclear shape, impacting gene accessibility and epigenetic regulation. Within the calcium signaling branch, calcineurin dephosphorylates NFATC family members (e.g., NFATC1, NFATC2, NFATC3), enabling their nuclear translocation. The R2482H variant can affect calcineurin’s sensitivity to Ca²⁺, thereby influencing NFATC-driven ECM gene expression and MMP activity. In addition to integrin and ion channel inputs, GPCR activation and primary cilia bending under mechanical load provide further layers of mechanosensing. Conformational shifts in GPCRs trigger heterotrimeric G proteins, reinforcing integrin–FAK and Ca²⁺ pathways. Primary cilia respond to fluid shear or matrix deformation by regulating intraflagellar transport, thereby modulating transcription factors including scleraxis (Scx), Mohawk (Mkx), and EGR1. Finally, mechanical signals converge at the transcriptional and epigenetic level, where factors such as Scx, Mkx, EGR1, and YAP/TAZ integrate cytoskeletal tension cues to control ECM-related gene programs. Epigenetic modifications (histone alterations, DNA methylation, microRNAs) refine collagen and MMP expression, while growth factors (TGF-β, FGF, CTGF) further reinforce fibril assembly and homeostatic tissue remodeling.
Mechanosensitive
ion channels—including members of the TRP
(Transient Receptor Potential) superfamily (e.g., TRPV4) and PIEZO
channels (e.g., PIEZO1)—translate physical deformation of the cell
membrane into ionic fluxes [93–95]. When stretched or subjected to
fluid shear, these channels open and permit rapid influx of ions,
predominantly Ca²⁺,
into the cytoplasm. Intracellular calcium elevation activates
calmodulin-dependent kinases or phosphatases (including calcineurin),
which then modulate a host of transcriptional regulators [96–98].
Increased cytosolic Ca²⁺ also impacts the Rho family of GTPases
(RhoA, Rac1, Cdc42), controlling actin stress-fiber formation and
focal adhesion organization [99–100]. These cytoskeletal
reconfigurations affect nuclear geometry and chromatin accessibility,
linking external mechanical cues to epigenetic modifications (e.g.,
histone acetylation, DNA methylation) and facilitating the
loading-specific regulation of ECM-related genes.
A key
component of the calcium-mediated branch of mechanotransduction in
ligament cells is calcineurin,
a Ca²⁺/calmodulin-dependent serine/threonine phosphatase that can
be referred to in some contexts as “calcineurin 3.” Calcineurin
dephosphorylates members of the NFAT
(Nuclear Factor of Activated T-cells) family—such as NFATC1,
NFATC2, and NFATC3—enabling their translocation to the nucleus
[96–98]. Once inside the nucleus, these NFATC factors orchestrate
the expression of ECM components, MMPs, and other remodeling proteins
critical for ligament integrity. The R2482H
variant, typically found in the regulatory subunit gene of
calcineurin (PPP3R1), can alter the phosphatase’s sensitivity to
Ca²⁺ levels, in turn affecting the magnitude or speed of NFATC
activation. Such a variant may predispose certain individuals to
differential ligament adaptations or altered healing outcomes,
exemplifying how genetic polymorphisms intersect with mechanical
loading to influence tissue phenotype [81–83]. Besides integrin and
ion channel signaling, ligament fibroblasts can detect mechanical
cues via GPCRs
[101–102]. Under load, G protein-coupled receptors undergo
conformational shifts that trigger heterotrimeric G proteins, adding
an extra layer of molecular crosstalk with integrin-FAK signals and
Ca²⁺-mediated pathways. Moreover, primary cilia—antenna-like
organelles comprising a microtubule core—may project from ligament
fibroblasts into the pericellular matrix [103–104]. Bending of
these cilia in response to fluid shear or ECM deformation modulates
intraflagellar transport, influencing downstream signaling modules
that converge on the nucleus to regulate transcription factors like
Scx, Mkx, and EGR1. Defects in ciliary assembly or function can thus
compromise ligament fibroblasts’ ability to sense and adapt to
changing mechanical environments.
Tensile loading predominantly arises from joint movements that elongate the ligament, creating strain on collagen fibrils and associated focal adhesions. This strain alters the conformation and clustering of integrin subunits (e.g., α5β1, αvβ3), triggering the recruitment of focal adhesion proteins such as talin, vinculin, paxillin, and α-actinin to adhesion complexes [87–89]. These nascent adhesions activate FAK and Src family kinases, converging on MAPK (ERK, JNK, p38) and PI3K/AKT pathways [30–32] [90–92]. ERK mediates proliferation and collagen gene expression, while JNK and p38 reorganize the actin cytoskeleton and regulate matrix metalloproteinases (MMPs). The PI3K/AKT pathway coordinates cell survival and metabolic adaptation, ensuring that collagen deposition and fibril alignment match the tensile demands imposed on the tissue.
By contrast, compressive loading is often transmitted through regions near entheses (bone–ligament junctions) or via changes in tissue hydrostatic pressure and fluid flow within the ECM. Under compression, ligamentocytes may experience altered osmotic balances that activate mechanosensitive ion channels such as TRPV4, which is sensitive to volumetric changes and hydrostatic pressure [93–95]. The resultant influx of Ca²⁺ engages calmodulin-dependent kinases or phosphatases (e.g., calcineurin), further modulating Rho GTPase activity (RhoA, Rac1, Cdc42) and promoting cytoskeletal reorganization. These shifts in cytoskeletal architecture can ultimately influence nuclear shape, chromatin accessibility, and the expression of ECM proteins vital for accommodating compressive stress.
Meanwhile, shear loading arises from fluid movement across cell surfaces or from sliding between collagen fibers under torsional stresses. Shear forces can open mechanosensitive ion channels, including PIEZO (PIEZO1) and certain TRP family members, permitting rapid ionic flux—primarily Ca²⁺—into the cytoplasm [93–95]. Elevated Ca²⁺ activates calcineurin, thereby regulating NFATC-dependent transcription and other Ca²⁺-responsive pathways [96–98]. In addition, shear stress can bend primary cilia, altering intraflagellar transport and triggering signaling cascades that converge on transcription factors like Scx, Mkx, and EGR1. Taken together, tensile, compressive, and shear inputs each engage overlapping yet distinct molecular circuits, enabling ligamentocytes to fine-tune ECM remodeling depending on the specific mechanical context.
At the transcriptional and epigenetic level, mechanical signals converge on factors such as Scx, Mkx, EGR1, and YAP/TAZ, each playing a distinct role in specifying ligament lineage and ECM gene expression [105–107]. Alterations in cytoskeletal tension may reshape the nuclear lamina and affect how these factors and their coactivators access DNA. Simultaneously, epigenetic mechanisms—including modifications of histone proteins (acetylation, methylation, phosphorylation), chromatin remodeling complexes, and the action of noncoding RNAs—further refine ligament gene regulation [108–109]. MicroRNAs (e.g., miR-29 family, miR-221) can dampen excessive collagen transcription or MMP production, optimizing matrix turnover [110–111]. Meanwhile, mechanical loading can liberate latent growth factors like TGF-β or modulate the activity of FGF and CTGF, reinforcing collagen fibril assembly and crosslinking in tandem with integrin/ion channel signals [39–41] [112]. Such growth factor signaling synchronizes fibroblast proliferation, ECM deposition, and fibril organization to maintain structural homeostasis or promote repair following injury [113]. Hence, mechanotransduction in ligament cells emerges as an integrated molecular circuit wherein integrins, mechanosensitive ion channels, GPCRs, and primary cilia provide multifaceted sensing capabilities, while calcineurin–NFATC signaling, Rho GTPases, transcription factors, and epigenetic regulators translate these signals into coordinated ECM remodeling. Under physiological loads, these pathways uphold balanced collagen synthesis, tissue resilience, and functional stability. However, perturbed mechanical environments—whether from insufficient or excessive loading, acute trauma, or genetic factors like R2482H—can yield disorganized collagen architectures, delayed healing, or degeneration.
Molecular Adaptations to Mechanical Loading
Ligament cells exhibit remarkable sensitivity to their mechanical environment, continually adjusting their molecular output in response to variations in load (Table 2). Under physiologic levels of cyclic tensile loading—mimicking normal joint motion—ligament fibroblasts maintain a tightly regulated equilibrium between ECM synthesis and degradation, ensuring that tissue remodeling aligns with functional demands [42–44, 114–116]. In this balanced state, integrin-mediated signaling and focal adhesion assembly foster robust cytoskeletal organization, sustaining the elongated, spindle-shaped morphology characteristic of healthy ligament cells. The resulting mechanical cues influence key transcription factors, including Scx and Mkx, which are central to specifying and preserving the ligamentous phenotype [45–48, 117–119]. By promoting the expression of collagen I and III, proteoglycans, and associated glycoproteins, physiologic loading reinforces tensile strength, elasticity, and structural integrity. At the molecular level, physiological loading also impacts the activity of signaling cascades such as the MAPK (ERK, JNK, p38) and PI3K/AKT pathways. These pathways converge on the nucleus, modulating transcription factor binding to gene promoters and enhancers, thereby enhancing the synthesis of ECM proteins that support tissue homeostasis [120–122]. Balanced loading ensures that the rates of collagen deposition, fibrillogenesis, and crosslinking match the tissue’s mechanical requirements. Through the controlled production of proteoglycans like decorin and biglycan, ligament cells maintain appropriate collagen fibril spacing, contributing to optimized viscoelastic properties that resist dynamic and complex loading patterns [123–124].

Table 2: Table delineates the molecular and cellular responses of ligament cells under different mechanical loading modalities: Physiologic Loading, Insufficient Loading, and Excessive Loading. This table highlights key aspects such as signaling pathways, gene expression, ECM composition, and clinical implications, supported by references
In contrast, deviations from this equilibrium—either by mechanical underloading or excessive strain—shift the molecular landscape toward a catabolic, degenerative state. Inadequate load (e.g., during immobilization) decreases integrin clustering and focal adhesion stability, reducing FAK phosphorylation and downstream signaling [125–126]. Without the requisite mechanical stimuli, ligament fibroblasts downregulate anabolic genes while upregulating catabolic mediators. These changes often manifest as an increase in collagen III at the expense of collagen I, disruption of fibril organization, and altered expression of tenascin-C, a glycoprotein that influences cell–matrix interactions and tissue repair processes [49–51, 127–129]. The result is a more disorganized ECM, reduced stiffness, and diminished load-bearing capability. Excessive mechanical loading, on the other hand, can induce cellular stress responses that involve the release of pro-inflammatory cytokines such as IL-1β and TNF-α [52–54, 130–132]. These cytokines stimulate pathways that enhance the synthesis and activation of matrix metalloproteinases (MMPs), enzymes that degrade collagen and other ECM components. As ECM integrity declines, the tissue becomes weaker and more susceptible to injury. Excessive loading may also alter the expression of key regulatory microRNAs that fine-tune collagen and MMP gene expression, reinforcing the breakdown of ECM and driving the tissue toward pathological remodeling [133–135].
In essence, the molecular adaptations of ligament cells to mechanical loading form a delicate continuum. At one end, physiologic loading maintains homeostasis by orchestrating a network of signaling molecules, transcription factors, and epigenetic regulators that promote collagen synthesis, appropriate proteoglycan levels, and stable cell phenotypes. At the other extreme, insufficient or excessive loading conditions lead to a downward spiral of altered integrin signaling, inflammatory mediator release, and enhanced ECM degradation.
Tendon-to-Ligament Transition. Anterior Cruciate Ligament Graft Remodeling
A
clinically relevant example of ligament cell plasticity at the
molecular and cellular levels is demonstrated after
anterior cruciale ligament (ACL) reconstruction with the use on
tendon grafts. Common donor tissues, such as the hamstring
(semitendinosus/gracilis tendon graft)
or patellar tendon, initially retain their inherent tendon-like
molecular profile, characterized by a highly aligned, predominantly
type I collagen matrix and minimal elastin or proteoglycan content
[55–57, 136–138]. Placed into the knee joint, these tendon grafts
are suddenly exposed to a complex mechanical milieu fundamentally
different from the unidirectional tensile environment of a tendon’s
native placement.
Over time, the graft undergoes a process termed “ligamentization,” in which mechanical loading patterns, local biochemical signals, and the joint’s synovial environment collaborate to drive a molecular and cellular transformation from a tendon-like to a ligament-like phenotype [58–61, 139–140]. This biological adaptation involves complex remodeling, including cellular infiltration, vascularization, extracellular matrix reorganization, and collagen maturation. Ligamentization occurs in distinct phases, beginning with early necrosis, followed by revascularization, cellular repopulation, and progressive structural integration into the knee joint. The tendon graft healing process in a bone tunnel has been studied in animal models [141]. Initially, the graft-tunnel interface fills with vascular granulation tissue rich in type III collagen. Vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF) stimulate macrophage influx and fibroblast enlargement. Chondroid cells deposit type II collagen, supporting new bone formation [141]. By 3–4 weeks, the matrix matures into Sharpey-like collagen fibers bridging the bone and graft. These fibers, composed of type III collagen, integrate into surrounding bone to resist shear stress, with their size correlating to graft pull-out strength. The process takes 8–30 weeks [141], with graft attachment strengthening as bone grows into the interface tissue [142].
At the heart of this transformation are changes in mechanotransduction pathways: integrins bind to newly abundant ECM components, including type III collagen, elastin, decorin, and biglycan. These integrin-mediated adhesions remodel focal adhesions and alter the actin cytoskeleton organization, shifting intracellular tension and triggering new signaling cascades [62–64, 143-144]. On the molecular level, this transition is orchestrated by shifts in gene expression that reflect the ligament’s need to accommodate more complex, multidirectional loads. Transcription factors such as Scx and Mkx may adjust their relative expression patterns in response to altered mechanical stress and growth factor gradients [145–147]. Additionally, extracellular signals from TGF-β, FGF, and VEGF support ECM remodeling, neo-vascularization, and cellular proliferation [148–150]. TGF-β, for instance, drives the production of type III collagen and elastin, while VEGF promotes angiogenesis, ensuring that the evolving graft is well-vascularized and metabolically supported. FGFs and CTGF further influence collagen fibrillogenesis and crosslinking, reinforcing the emerging ligamentous structure. Integrin signaling feeds into MAPK and PI3K/AKT pathways, modulating the balance of anabolic and catabolic genes. As these pathways are tweaked by the new mechanical environment, cells increase the synthesis of small leucine-rich proteoglycans (SLRPs) like decorin and biglycan [151–153]. These SLRPs guide collagen fibrillogenesis, adjusting fibril diameter distributions and introducing a characteristic crimp pattern more akin to ligament tissue. Over time, the mechanical properties evolve: the graft’s collagen fibrils become less linear and more varied in diameter and crimp, imparting increased compliance and an ability to handle shear and rotational forces typical of ligament loading [65–67, 154–156]. At the cellular level, epigenetic modifications (e.g., changes in histone acetylation, DNA methylation, and microRNA profiles) may also play a role in stabilizing the ligament-like transcriptional program [157–159]. These epigenetic changes help “lock in” the new molecular identity, ensuring that the once tendon-specific fibroblasts now consistently express genes associated with ligament biology and maintain a ligamentous ECM architecture.
Biologic augmentation strategies, such as delivering specific growth factors (e.g., TGF-β or VEGF), using gene editing techniques to upregulate ligament-associated transcription factors, or modulating epigenetic regulators, can further hasten the tendon-to-ligament transition [160–165].
Regulatory Networks and Transcriptional Control
Ligament cell fate and ECM composition are governed by an intricate regulatory landscape that integrates mechanical cues with transcriptional and epigenetic mechanisms. Central to this network are key transcription factors (TFs) that respond to the tissue’s mechanical environment and orchestrate the gene expression patterns needed for proper ligament structure and function. Scx, a basic helix-loop-helix transcription factor, is one of the most extensively studied regulators of tendon and ligament development. Scx directs the transcription of collagen genes (e.g., COL1A1, COL3A1) and other ECM components, ensuring the formation of robust, load-bearing collagen fibrils [166–168]. By interacting with cofactors and binding to enhancer regions of ECM-related genes, Scx helps maintain the ligament’s tensile strength and prevents the adoption of a non-ligamentous phenotype. Similarly, Mkx, a homeodomain transcription factor, plays critical roles in specifying tendon/ligament lineage commitment and sustaining ligament cell identity [169–170]. Mkx influences collagen fibrillogenesis and ECM organization, complementing the actions of Scx and ensuring that mechanical signals translate into a stable gene expression program. Another important transcription factor, EGR1, modulates the tissue’s response to injury and mechanical stress [171]. EGR1 is rapidly induced by extracellular signals and biomechanical loading, influencing genes involved in ECM remodeling, growth factor signaling, and cytoskeletal architecture. By adjusting the levels of EGR1, ligament cells can tune their repair strategies—upregulating MMPs and proteoglycans as needed during healing and downregulating them once homeostasis is restored. In this manner, EGR1 acts as a molecular rheostat that balances anabolic and catabolic processes in response to changing mechanical environments.
These transcription factors do not act in isolation. Instead, they function within larger regulatory networks where mechanical signals—transduced via integrins, focal adhesion complexes, and the cytoskeleton—converge on the cell nucleus to modulate TF activity [172–173]. Mechanical stretch or compression can alter TF nuclear localization, protein stability, and DNA-binding affinity. Certain mechanosensitive co-activators, such as YAP/TAZ, may also participate in this regulatory matrix [174–175]. YAP/TAZ activity is influenced by cytoskeletal tension and focal adhesion assembly, allowing cells to integrate mechanical information with biochemical cues and adjust gene expression patterns accordingly. In addition to these TF-driven regulatory circuits, epigenetic mechanisms provide another layer of fine-tuning. DNA methylation and histone modifications (e.g., acetylation, methylation) can dynamically alter chromatin structure, influencing the accessibility of transcription factor binding sites and thereby regulating the transcriptional output in ligament fibroblasts [176–178]. Mechanical cues have been shown to modulate chromatin organization, potentially through mechano-sensitive chromatin remodeling complexes that determine which genes are actively transcribed or repressed in response to load [179–180].
MicroRNAs (miRNAs) are emerging as critical post-transcriptional regulators of gene expression in ligament biology. These small, non-coding RNAs bind to target mRNAs and either repress their translation or induce their degradation. For instance, miRNAs can finely adjust collagen and MMP expression, ensuring that ECM synthesis and degradation remain in balance [181–183]. By selectively modulating levels of key structural proteins and enzymes, miRNAs help maintain tissue integrity and responsiveness to mechanical stimuli. Changes in miRNA expression profiles after injury or altered loading conditions can tilt the balance between anabolic and catabolic activities, contributing to either successful healing or degenerative changes. For example, delivering small molecules or gene-editing reagents (e.g., CRISPR/Cas9) could modulate the activity of Scx, Mkx, or EGR1, accelerating the tendon-to-ligament transition in ACL grafts [184–186]. Likewise, manipulating miRNA expression or histone-modifying enzymes could prevent unwanted fibrocartilaginous transformations, maintain ECM homeostasis, and promote superior mechanical function [187–188].
In essence, the molecular and cellular regulatory networks controlling ligament cell fate and ECM composition form an adaptive system that continually integrates mechanical and biochemical signals.
Growth Factors, Cytokines, and Inflammatory Mediators
Beyond structural proteins, the mechanical environment of the ligament coordinates a multifaceted molecular dialogue involving growth factors, cytokines, and other signaling molecules that guide ligament cell fate and ECM organization. Ligament fibroblasts—attuned to subtle changes in tissue tension and stiffness—respond to appropriate tensile strain by adjusting their secretome and receptor expression patterns [189–191]. When integrins at the cell surface detect physiological loading, they cluster within focal adhesions and engage intracellular adaptors to activate downstream signaling pathways [192–194]. This mechanotransduction process triggers the upregulation and release of key growth factors, such as TGF-β isoforms, which translocate to the nucleus via SMAD-dependent pathways. There, they influence chromatin structure and transcription factor binding, increasing the expression of collagen (particularly types I and III) and proteoglycans (such as decorin and biglycan). These ECM components enhance the tissue’s tensile strength, maintain proper fibril spacing, and preserve its viscoelastic properties [76, 195–197] Similarly, FGF and IGF-1 (Insulin-like Growth Factor 1) operate as potent modulators of cellular proliferation and ECM turnover by engaging their receptor tyrosine kinases and activating MAPK and PI3K/AKT cascades [198–200]. At the molecular level, these pathways converge on transcriptional regulators that orchestrate gene programs for ECM synthesis, cytoskeletal remodeling, and metabolic adaptations. FGF-driven signals, for instance, can fine-tune actin filament organization, influencing the spatial arrangement of collagen fibrils. IGF-1, in turn, can bolster anabolic metabolism, ensuring that fibroblasts have sufficient energy and substrates for ECM protein synthesis [201–203]. CTGF, another crucial factor, integrates mechanical cues and growth factor signals by localizing to focal adhesions and influencing collagen fiber alignment. CTGF’s presence refines fibril architecture, ensuring that newly deposited collagen fibers form robust, mechanically optimized networks [77–79, 204–206]. On a molecular scale, these growth factors modulate the balance between collagen crosslinking enzymes (e.g., lysyl oxidase) and proteolytic enzymes, guiding fibril maturation and stiffness. However, this delicately balanced molecular system can be perturbed by injury, abnormal loading patterns, or local hypoxia [207–209]. Excessive mechanical strain or reduced loading leads to alterations in integrin-mediated signaling, causing shifts in the fibroblast secretome. Under these conditions, pro-inflammatory cytokines such as IL-1β and TNF-α surge [210–212]. At the cellular level, these cytokines activate NF-κB and related transcriptional networks, upregulating MMPs and aggrecanases that degrade ECM components and weaken the overall scaffold [80–82, 213–215]. High levels of NO (Nitric Oxide) further compound these issues by disrupting growth factor receptor function and interfering with proper ECM protein folding and crosslinking [216–218]. NO’s reactive intermediates can modify the redox state of critical signaling proteins, diminishing the efficiency of growth factor-driven anabolic pathways and exacerbating collagen breakdown. This molecular environment fosters a feedback loop where degraded ECM fragments and persistent inflammatory signals perpetuate a cycle of tissue degeneration, scar formation, and biomechanical insufficiency.
From a cellular perspective, fibroblasts exposed to these pathological cues alter their gene expression profiles, increasing expression of stress-responsive genes and proteolytic enzymes while reducing collagen synthesis [219–221]. Changes in actin cytoskeleton organization and nuclear shape reflect shifts in transcription factor localization and chromatin accessibility, ultimately skewing the cell toward a catabolic phenotype [222–224]. Over time, these molecular changes reshape the local microenvironment, favoring fibrocartilaginous or scar-like tissue rather than a functional ligament structure. To reverse or prevent these maladaptive processes, interventions that target molecular pathways hold promise [225–227]. Such precise mechanical stimulation preserves TGF-β, FGF, IGF-1, and CTGF’s anabolic effects on collagen alignment and ECM deposition while dampening pro-inflammatory signals. Pharmacological agents or biologics that block IL-1β or TNF-α receptors, neutralize NO, or inhibit MMP activity can restore a more regenerative molecular environment. Gene editing tools or miRNA modulators could be deployed to stabilize anabolic gene expression patterns or to rescue compromised integrin and growth factor signaling pathways [228–230].
In summary, the molecular and cellular interplay between mechanical forces, growth factors, and inflammatory mediators is central to ligament homeostasis and repair. By understanding the precise molecular underpinnings—how integrin signaling interfaces with growth factor receptor activation, how cytokines distort transcriptional and epigenetic landscapes, and how NO modulates receptor sensitivity—clinicians and researchers can design strategies that maintain molecular homeostasis. This will improve ECM quality, ensure proper collagen fibrillogenesis, and ultimately lead to more resilient ligaments better able to withstand functional demands and recover from injury.
Stress Shielding in Ligaments
The concept of stress shielding in ligaments emerges when portions of the tissue are effectively protected or “shielded” from the mechanical strains they would normally experience (Fig. 2). Under healthy conditions, ligaments and tendons rely on tension and mechanical loading cues to maintain their structural integrity, ensuring proper cell alignment, collagen fibrillogenesis, and tissue homeostasis. However, when stress is diverted or reduced—such as by the presence of an adjacent stiffer material, a parallel healthy tissue segment, or changes in tissue composition—ligament cells are deprived of the mechanical signals needed to sustain normal ECM organization and tenogenic (ligament-like) gene expression [231–233]. Ligaments exhibit viscoelastic properties characterized by creep, hysteresis, and stress-relaxation—the phenomenon where tissue stress decreases over time under a constant strain. Normally, small, cyclic load-unload patterns and brief stress-relaxation intervals help maintain ligament health, allowing collagen fibrils to reorganize, water and solute to redistribute, and cells to recover metabolically. When stress is permanently shielded from certain ligament regions, those areas experience drastically reduced load and, thus, diminished opportunities for periodic stress-relaxation. Without adequate stress-relaxation cycles, fibroblasts fail to engage in the micro-adjustments (e.g., cytoskeletal remodeling, local collagen reorientation) vital for maintaining or restoring tissue tensile properties.
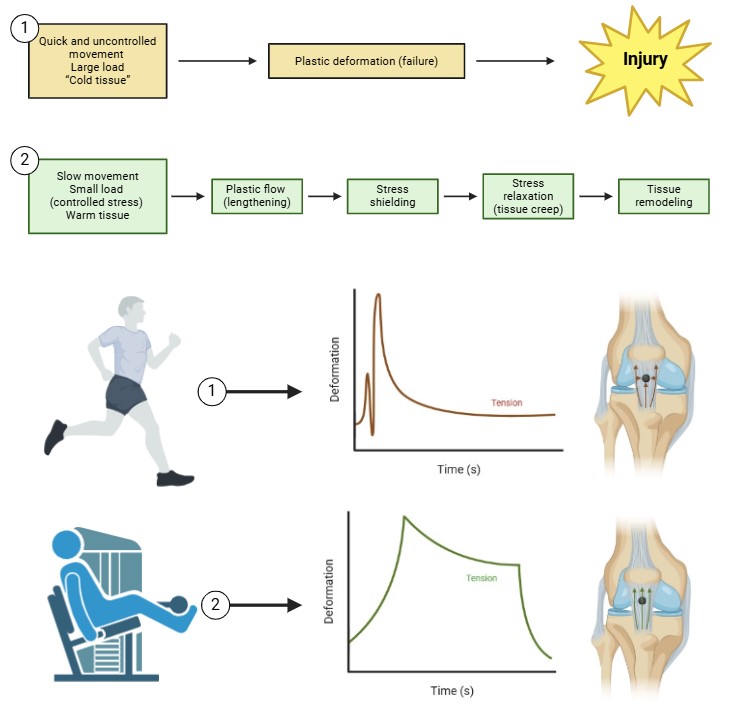
Fig. 2: This figure compares two loading scenarios—(1) quick, uncontrolled movement with large loads on “cold” tissues versus (2) slow, controlled movement with smaller loads on “warm” tissues—and illustrates how stress shielding and stress-relaxation can either contribute to injury or promote tissue remodeling. In the top panel (1), a sudden high load induces plastic deformation (failure) in the ligament, potentially leading to injury. Because the tissue is not given time to accommodate or reorganize collagen fibrils, the deformation overshoots the ligament’s elastic capacity, resulting in permanent damage. By contrast, the bottom panel (2) shows a gradual, lower-intensity load that proceeds through phases of plastic flow, stress shielding, stress relaxation, and ultimately tissue remodeling. Here, the tissue benefits from slow loading (e.g., warm-up exercises or controlled rehabilitation protocols) that facilitate creep, water redistribution, and cytoskeletal adjustments. Although stress shielding can still occur if certain regions of the ligament bear less load than others, timely stress-relaxation intervals allow fibroblasts to reorient collagen fibrils and maintain tenogenic gene expression. This is critical for avoiding disorganized, scar-like tissue and instead promoting healthier ECM architecture over time. The time–deformation graphs depict how, under quick heavy loads, tissue stress initially spikes and may quickly exceed its failure threshold. Under slower, methodical loading, tension climbs more gradually and then partially relaxes, allowing collagen realignment and micro-adjustments that protect ligament integrity. The knee joint schematic on the right underscores how these loading patterns impact ligament homeostasis: uncontrolled movement risks plastic deformation and injury, whereas controlled stress cycles combined with adequate rest and stress-relaxation intervals encourage adaptive remodeling and strengthen the ligament’s load-bearing capacity. High intensity activities monitoting is crucial in injury prevention.
In healing or injured ligament tissues, the formation of a compliant, disorganized scar in parallel with stiffer, healthier tissue leads to a significant mismatch in mechanical properties [234–235]. Because the intact portion of the ligament (or an external stiff structure, like a surgical implant) bears most of the load, the more compliant scar region undergoes relatively less tension. This reduction in stress transmission to the scar is known as stress shielding. The under-loaded scar tissue experiences minimal stress-relaxation episodes, limiting its ability to reorganize collagen fibrils. Ligament fibroblasts in the shielded region lose crucial mechanical cues, reduce matrix turnover, and fail to form well-aligned collagen fascicles capable of bearing load [236–238]. Without adequate tensile loading, cells within the scar cannot properly align; consequently, collagen fibrils remain disordered and fail to mature into a load-bearing, ligament-like matrix. Over time, diminished stress-relaxation intervals can further hasten the fibrocartilaginous transition of the tissue, making it more vulnerable to mechanical failure.
Molecularly, in the absence of sufficient tensile stress and normal stress-relaxation cycles, transcription factors such as Mohawk (Mkx) and Egr1 remain low, while chondrogenic markers like Sox9 and Col2a1 rise [239–241]. In a properly stressed ligament, periodic strain (and associated stress-relaxation) triggers integrin–FAK–MAPK signaling that drives collagen type I expression, organizes fibrils, and promotes the tenogenic phenotype. By contrast, shielded regions fail to sustain these signals. Elevated Sox9, collagen type II, and proteoglycan content emerges, resembling tissue more akin to cartilage than ligament. Scar tissue retains immature collagen crosslinks and smaller fibril diameters, lacking the robust load-bearing capacity typical of healthy ligament [242–244]. Experimental evidence supports the role of stress shielding in this detrimental cascade. Insertion of a stiffer material in parallel with a healthy tendon or ligament decreases the tensile forces experienced by the native tissue, leading to decreased load-bearing capacity, collagen disorganization, and hypercellularity—changes characteristic of scar formation [245–247]. These shielded regions also exhibit insufficient stress-relaxation behavior, indicating minimal opportunities for reorienting collagen fibrils and remodeling the matrix.
Conversely, lowering the stiffness of the healthy portion of the tissue or adjusting loading protocols (such as introducing isometric tension that facilitates creep and thus better load transfer to scarred areas) can reintroduce mechanical signals to shielded cells [248–250]. These signals restore partial stress-relaxation cycles and help fibroblasts regain a tenogenic phenotype. Elevated Mkx/Egr1 expression, improved collagen alignment, and more mature crosslinking patterns are often observed when mechanical cues are reestablished.
In summary, stress shielding in ligaments is a mechanically driven phenomenon wherein portions of a healing or regenerating ligament experience insufficient tensile loading and inadequate stress-relaxation intervals. This lack of mechanical input prevents proper ECM maturation and gene expression patterns essential for restoring healthy ligament structure and function, thereby contributing to the persistence of disorganized, scar-like tissue rather than promoting regenerative healing. Reintroducing tension—via modifications in loading protocols, stiffness gradients, or physical rehabilitation regimens that encourage periodic stress-relaxation—may potentially restore normal collagen architecture and bolster the mechanical resilience of the ligament over time.
ACL Microtrauma and Overuse
Microtrauma accumulates at the molecular and cellular levels as the ACL is subjected to repetitive, sub-failure loads that exceed its capacity for timely repair. At the core of this process is the ligament’s resident fibroblasts, which continuously sense and respond to mechanical signals through integrin-mediated focal adhesions, the cytoskeleton, and associated mechanotransduction pathways [251–253]. Under healthy loading conditions, these cells maintain the ECM by synthesizing collagen and other proteins, adjusting fibril diameter and organization, and balancing anabolic and catabolic activities. However, when microtrauma occurs faster than the tissue can repair, molecular homeostasis is disrupted (Fig. 3).
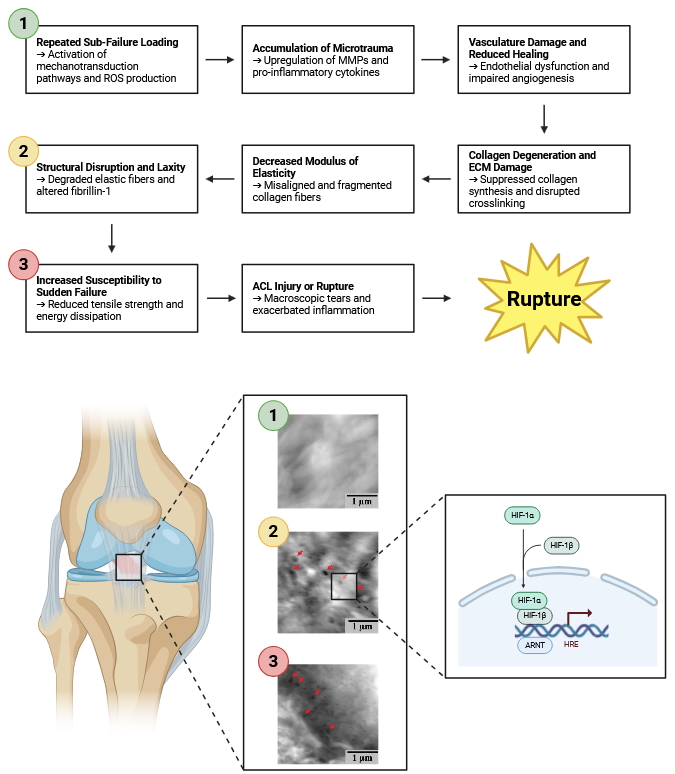
Fig. 3: Presents a detailed schematic divided into eight interconnected panels, each representing a distinct stage in the progression from repeated sub-failure loading to ACL injury or rupture. The sequence begins on the left with Repeated Sub-Failure Loading and culminates on the right with ACL Injury or Rupture. Each panel not only depicts the mechanical and structural alterations within the ligament but also delves into the underlying molecular responses that drive these changes. The progression commences with Repeated Sub-Failure Loading, where athletes engage in repetitive movements such as jumping, cutting, and pivoting. These activities subject the ACL to cyclic tensile and shear forces that are insufficient to cause immediate structural failure but are significant enough to activate mechanotransduction pathways within ACL fibroblasts. This mechanical stress triggers the activation of transcription factors NF-κB (Nuclear Factor kappa-light-chain-enhancer of activated B cells) and AP-1 (Activator Protein 1), which translocate to the nucleus to initiate the expression of various inflammatory genes. Concurrently, the production of Reactive Oxygen Species (ROS) increases within ligament cells, symbolizing the onset of oxidative stress. As loading continues, Accumulation of Microtrauma occurs, evidenced by minor, reversible damage to the extracellular matrix (ECM), particularly the collagen fibers. At the molecular level, there is an elevation in Matrix Metalloproteinases (MMP-1 and MMP-13), enzymes responsible for degrading collagen and other ECM proteins. The synthesis of pro-inflammatory cytokines such as Interleukin-1 beta (IL-1β) and Tumor Necrosis Factor-alpha (TNF-α) is upregulated, enhancing MMP activity while simultaneously suppressing ECM synthesis. Additionally, alterations in integrin signaling pathways disrupt the critical communication between cells and the ECM, further exacerbating tissue degradation. Progressing to Vasculature Damage & Reduced Healing, the persistent microtrauma inflicts damage on the microvasculature within the ACL, leading to reduced blood flow. This hypoxic environment stabilizes Hypoxia-Inducible Factor 1-alpha (HIF-1α), which alters gene expression to adapt to low oxygen conditions. However, chronic stabilization of HIF-1α results in maladaptive responses, including the downregulation of Vascular Endothelial Growth Factor (VEGF), thereby inhibiting angiogenesis. Concurrently, apoptotic pathways are activated in ACL fibroblasts through the involvement of caspases and the regulation of BAX/Bcl-2 proteins, leading to programmed cell death and further diminishing the ligament's capacity for repair and regeneration. The subsequent stage, Collagen Degeneration & ECM Damage, is marked by diminished and disorganized collagen fibers within the ligament. There is a suppression of type I collagen genes (COL1A1 and COL1A2) and a downregulation of lysyl oxidase, an enzyme crucial for collagen crosslinking. This weakening of collagen crosslinks undermines the structural integrity of the collagen fibers. Additionally, aggrecanases degrade glycosaminoglycans (GAGs), essential components that maintain the ECM's viscoelastic properties. The degradation of GAGs further compromises the ligament's structural resilience, making it more susceptible to mechanical deformation. Moving to Decreased Modulus of Elasticity, the ligament exhibits reduced stiffness and increased flexibility. This change is a direct consequence of misaligned and fragmented collagen fibers, which disrupt the uniform distribution of mechanical stress across the ligament. Oxidative modifications, such as carbonylation, impair the mechanical properties of collagen, while a reduced density of intact collagen fibrils diminishes the ligament’s elastic modulus. These molecular alterations weaken the ligament's ability to absorb and distribute tensile forces effectively. In the stage of Structural Disruption & Laxity, the ligament appears stretched and less taut, indicating increased laxity. Elastin fibers undergo enzymatic degradation, and the expression of tenascin-C, a glycoprotein involved in tissue remodeling, is upregulated. These changes contribute to the disorganization of the ECM, further compromising the ligament's structural integrity. The loss of fibril alignment adversely affects load distribution, rendering the ligament more prone to deformation and injury during normal activities. As damage accumulates, the ligament reaches a state of Increased Susceptibility to Sudden Failure. The overall structure of the ACL is significantly weakened, with visible areas of potential rupture. Cellular viability decreases due to ongoing apoptosis and necrosis of ACL fibroblasts, impairing the ligament’s ability to maintain and repair the ECM. Disruptions in integrin and growth factor signaling hinder the ligament’s adaptive responses to mechanical stress, while protein misfolding and aggregation further compromise cellular functions. These molecular dysfunctions collectively reduce the ligament's resilience, making it highly susceptible to sudden failure. The final panel, ACL Injury or Rupture, depicts the complete tear of the ACL accompanied by surrounding inflammation. The collagen matrix can no longer withstand the tensile forces, resulting in macroscopic tears. An inflammatory cascade is activated, releasing damage-associated molecular patterns (DAMPs) that exacerbate tissue injury. The body's attempt to heal the damaged ligament involves scar tissue formation, characterized by disorganized fibroplasia that fails to restore the original function and integrity of the ligament. Figure adapted from Chen J et. al., 2019).
Molecularly, repetitive sub-failure strains can cause partial unfolding of collagen’s triple-helical structure at the fibrillar and sub-fibrillar levels. This mechanical deformation exposes cryptic binding sites that increase susceptibility to enzymatic cleavage by MMPs and aggrecanases [254–256]. The altered mechanical environment shifts fibroblast gene expression patterns, downregulating genes associated with stable collagen assembly and upregulating those involved in ECM remodeling and degradation. Over time, these changes compromise the uniform alignment and crosslinking of collagen fibrils. As collagen fibrils become disorganized or partially denatured, the mechanical stiffness and tensile strength of the ligament decrease, lengthening the force-displacement “toe region” of the stress-strain curve and increasing ligament laxity [257–259].
At the cellular level, the reduced ECM integrity and altered load distribution modify integrin engagement, focal adhesion assembly, and downstream signaling via MAPKs, PI3K/AKT, and Rho GTPases [260–262]. Fibroblasts exposed to these altered mechanical cues may produce more inflammatory mediators and shift toward a more catabolic phenotype, increasing MMP secretion and decreasing collagen synthesis. In addition, vasculature damage from microtrauma diminishes nutrient and oxygen delivery, inducing localized hypoxia [263–265]. Ligament cells, possessing relatively low baseline metabolic activity, are generally resistant to short-term hypoxia. However, chronic or repeated episodes can hamper cellular respiration, reduce ATP production, and limit the resources needed for collagen synthesis and proper fibril maturation. The resulting ischemic environment may trigger oxidative stress and increase reactive oxygen species (ROS) production, further impacting protein folding, crosslink integrity, and cell membrane stability [266–268]. Cellular responses to microtrauma also involve subtle changes in mechanosensitive ion channels, growth factor receptor signaling, and epigenetic regulation [269–271]. For instance, changes in loading frequency or magnitude can alter the expression of transcription factors like Scx and Mkx, as well as microRNAs controlling collagen deposition and MMP activity. Poor vascularity within certain regions of the ACL, especially at the entheses, slows the infiltration of progenitor cells and reparative growth factors, prolonging the duration during which fibroblasts operate under suboptimal conditions [272–274]. This spatial heterogeneity in blood supply can create local “hotspots” where microtrauma accumulates, giving rise to region-specific ECM degeneration and eventual mechanical failure.
In addition, excessive exercise intensity or poor recovery strategies can chronically elevate ROS levels and pro-inflammatory cytokines in the ligament microenvironment (Fig. 4). This metabolic and oxidative imbalance places further strain on fibroblasts’ repair capacity, reducing their ability to maintain a stable collagen network. Intermittent exercise, with adequate rest and nutrient support, can improve redox homeostasis and antioxidant defenses, enabling fibroblasts to better manage oxidative stress and resume anabolic activities [275–277]. Appropriate timing of collagen peptide supplementation and balanced amino acid availability may support fibroblast function, enhancing collagen synthesis and improving the ECM’s resilience to repeated loading [278–279]. Ultimately, microtrauma-induced changes at the molecular and cellular levels diminish the ligament’s modulus of elasticity and predispose it to sudden fatigue failure under nominal loads. Repetitive sub-failure strain compromises the delicate equilibrium between ECM deposition and degradation, shifts fibroblast phenotypes, and disrupts nutrient and oxygen delivery, collectively weakening the ACL’s structural integrity [280–282]. By optimizing cellular metabolism, redox homeostasis, and growth factor signaling, it may be possible to enhance the ACL’s capacity for microtrauma repair, thereby extending its functional endpoint and reducing the risk of catastrophic rupture. In cases where surgical intervention is not immediately required or contraindicated, conservative treatment strategies play a crucial role in managing ACL injuries. Structured rehabilitation programs focusing on neuromuscular training, proprioception enhancement, and quadriceps-hamstring coactivation can aid in stabilizing the knee joint and compensating for ligamentous insufficiency. Additionally, interventions such as bracing, controlled progressive loading, and biomechanical gait retraining help mitigate excessive stress on the ACL while promoting adaptive tissue remodeling.
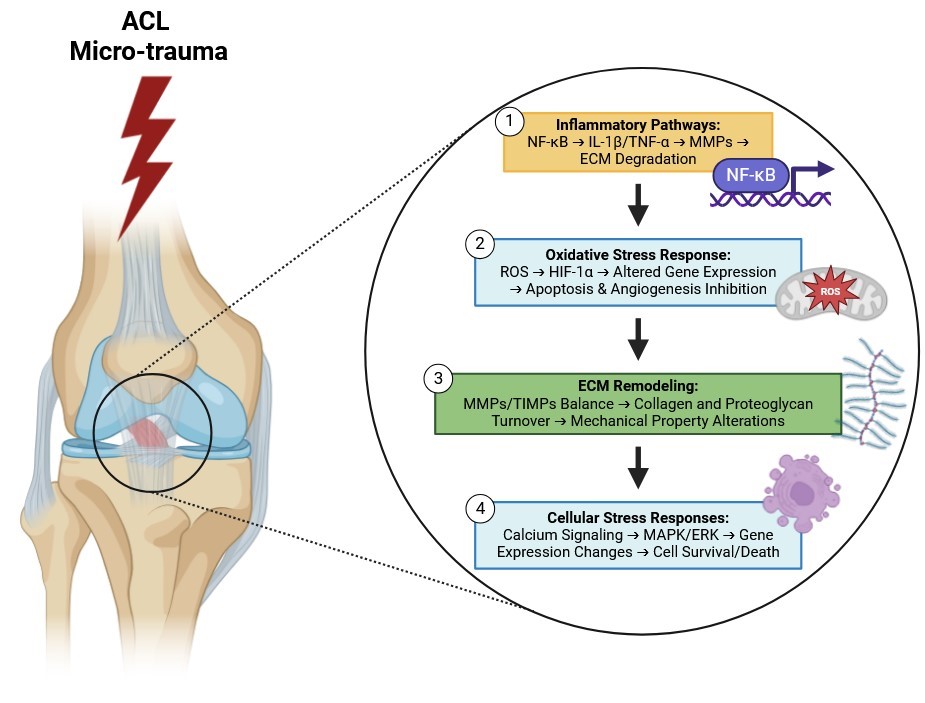
Fig. 4: Presents a multi-layered schematic that delineates the intricate molecular mechanisms driving the progression from repeated sub-failure loading to ACL injury or rupture. The illustration is organized into four primary interconnected pathways—Inflammatory Pathways, Oxidative Stress Response, Extracellular Matrix (ECM) Remodeling, and Cellular Stress Responses—each highlighting a critical aspect of the cellular and molecular responses to mechanical stress within the ACL. These pathways collectively demonstrate the complex interplay between inflammatory signaling, oxidative stress, ECM degradation, and cellular stress responses, culminating in the structural failure of the ligament. The progression begins with repeated sub-failure mechanical loading, where cyclic tensile and shear forces are applied to ACL fibroblasts without causing immediate structural failure. This mechanical stress activates the NF-κB (Nuclear Factor kappa-light-chain-enhancer of activated B cells) signaling pathway through mechanotransduction mechanisms. Once activated, NF-κB translocates to the nucleus, initiating the expression of various inflammatory genes. This upregulation leads to the synthesis of pro-inflammatory cytokines such as Interleukin-1 beta (IL-1β) and Tumor Necrosis Factor-alpha (TNF-α), which amplify the inflammatory response within the ligament tissue. These cytokines further stimulate the expression and activation of Matrix Metalloproteinases (MMPs), including MMP-1 and MMP-13, enzymes responsible for degrading key components of the ECM, particularly type I collagen. The heightened activity of MMPs disrupts the integrity of the ECM, thereby compromising the ACL's ability to withstand tensile forces and contributing to ligament laxity and vulnerability to injury. Simultaneously, sub-failure mechanical loading elevates the production of Reactive Oxygen Species (ROS) within ACL fibroblasts, inducing oxidative stress. Elevated ROS levels, coupled with resulting hypoxic conditions, stabilize Hypoxia-Inducible Factor 1-alpha (HIF-1α) by preventing its degradation. Stabilized HIF-1α translocates to the nucleus and alters gene expression to adapt to low oxygen environments. However, chronic stabilization of HIF-1α leads to maladaptive responses, including the induction of apoptotic signaling pathways involving caspases and the regulation of BAX/Bcl-2 proteins, culminating in programmed cell death of ACL fibroblasts. Additionally, chronic inflammation and HIF-1α-mediated gene expression downregulate Vascular Endothelial Growth Factor (VEGF), impairing the formation of new blood vessels and further reducing the ACL's capacity for repair and regeneration. In the realm of Extracellular Matrix (ECM) Remodeling, the balance between Matrix Metalloproteinases (MMPs) and their natural inhibitors, Tissue Inhibitors of Metalloproteinases (TIMPs), is crucial. Chronic mechanical stress disrupts this balance by upregulating MMPs while downregulating TIMPs, tipping the scale toward ECM degradation. Enhanced MMP activity leads to increased breakdown of type I collagen and proteoglycans such as decorin and biglycan, which are essential for maintaining the viscoelastic properties of the ECM. The degradation of these components results in reduced tensile strength and elasticity of the ACL, making the ligament more susceptible to deformation and injury under normal physiological loads. Concurrently, Cellular Stress Responses are triggered by repetitive mechanical loading, which induces transient increases in intracellular calcium levels within ACL fibroblasts. Elevated calcium acts as a secondary messenger, initiating various intracellular signaling cascades, including the activation of the Mitogen-Activated Protein Kinase (MAPK) and Extracellular Signal-Regulated Kinase (ERK) pathways. These pathways regulate a wide array of cellular processes, including proliferation, differentiation, and apoptosis. Activation of MAPK/ERK leads to changes in the expression of genes involved in cell survival, proliferation, and ECM production. However, chronic activation under persistent mechanical stress can shift the balance toward catabolic gene expression profiles, resulting in decreased cellular viability. Depending on the context and duration of signaling, ACL fibroblasts may undergo apoptosis or necrosis, thereby impairing the ligament’s capacity to maintain and repair the ECM. The integration of these four pathways—Inflammatory Pathways, Oxidative Stress Response, ECM Remodeling, and Cellular Stress Responses—creates a synergistic network that drives the degeneration of the ACL. Inflammatory signaling exacerbates ECM degradation by increasing MMP activity, while oxidative stress impairs vascular health and promotes apoptosis of essential fibroblasts. The imbalance in ECM remodeling further weakens the ligament’s structural integrity, and the cellular stress responses reduce the ligament’s ability to repair and regenerate damaged tissue. This cumulative effect of molecular and structural changes ultimately leads to the rupture of the ACL, especially when the ligament is subjected to additional mechanical loads that exceed its compromised capacity.
Ligament Chronobiology: Loading and Unloading
Ligaments exist in a dynamic mechanical environment where the interplay of loading and unloading cycles orchestrates their molecular and cellular behavior. Chronobiology—the time-dependent patterning of biological processes—provides a framework for understanding how the frequency, duration, and intensity of mechanical stimuli modulate the ECM composition, collagen synthesis, and cellular phenotype of ligament fibroblasts over periods of strain and rest [283–285]. Rather than a static tissue, the ligament responds to mechanical cues in rhythmic fluctuations that shape gene expression, protein turnover, and cellular metabolism. At the molecular level, integrin-mediated mechanotransduction pathways continuously gauge tensile forces. During loading phases, integrin engagement with collagen and proteoglycans stabilizes focal adhesions, activating FAK and downstream MAPK (ERK, JNK, p38) and PI3K/AKT cascades [286–288]. This activation amplifies signals that promote collagen gene transcription (COL1A1, COL3A1), ECM deposition, and proper fibril alignment. Mechanosensitive ion channels, such as TRP and PIEZO channels, open under tensile or shear forces, allowing Ca²⁺ influx [289–291]. Elevated intracellular Ca²⁺ fine-tunes cytoskeletal organization and enhances transcription factor activity, reinforcing ECM assembly. Growth factors like TGF-β, FGF, and CTGF integrate into these pathways, leveraging mechanical cues to boost collagen crosslinking, fibrillogenesis, and mechanical strength [292–294].
Conversely, unloading intervals modulate these molecular networks differently. Without sufficient mechanical stimulation, integrin clustering diminishes, lowering FAK activity and reducing anabolic signaling [295–297]. Closed mechanosensitive ion channels limit Ca²⁺ entry, diminishing transcription of ECM proteins and potentially upregulating catabolic mediators such as MMPs. With persistent unloading, fibroblasts may shift their gene expression patterns toward more fibrocartilaginous characteristics, reducing collagen tensile strength and increasing tissue vulnerability [298–300]. Epigenetic modifications (e.g., histone acetylation, DNA methylation) and altered growth factor signaling further entrench these changes, leading to disorganized collagen fibrils and weaker mechanical properties [301–303]. Ligament fibroblasts also exhibit circadian clock genes, including BMAL1 and CLOCK, which form the core transcriptional and translational feedback loops that drive rhythmic gene expression [283–285]. Mechanical stimuli can synchronize these circadian regulators, influencing downstream targets like PER and CRY, which in turn modulate ECM metabolism, MMP activity, and collagen homeostasis. During loading phases aligned with the circadian rhythm, BMAL1/CLOCK complexes may reinforce signals from FAK or Ca²⁺-dependent cascades, ensuring optimal collagen synthesis and repair. By contrast, misalignment—such as prolonged unloading or irregular mechanical stress—could disrupt the normal oscillations of BMAL1/CLOCK, attenuating the anabolic signals and predisposing the ligament to collagen disorganization or slow healing.
At the cellular level, ligament fibroblasts interpret loading cycles through cytoskeletal adjustments and changes in nuclear morphology. During repeated loading, the cytoskeleton aligns stress fibers, improves actin-myosin tension, and maintains nuclear shape favorable for activating pro-tenogenic transcription factors like Scx and Mkx [304–306]. These patterns ensure balanced collagen turnover and steady ECM renewal. Appropriate unloading intervals allow cells to restore ATP and nutrient levels, recover from oxidative stress, and recalibrate growth factor signaling [307–309]. However, excessive unloading intervals or irregular load withdrawal can induce a gradual loss of tenogenic phenotype, undermining tissue integrity and stiffness [310–312].
Incorporating periodic rest phases between loading cycles provides opportunities for fibroblasts to upregulate antioxidant defenses, maintain mitochondrial function, and ensure a sustainable balance between reactive ROS production and detoxification [313–315]. This cyclical nature of mechanical input and metabolic restoration exemplifies the principle that ligaments thrive under conditions where loading and recovery follow a coordinated temporal pattern—one that also interfaces with circadian regulators like BMAL1 and CLOCK. These circadian mechanisms may help schedule ECM synthesis and degrade misfolded proteins in sync with daily fluctuations in mechanical activity, promoting both structural integrity and metabolic efficiency.
These insights emphasize that ligament healing and adaptation are governed by cycles of mechanical stimulation rather than a simple progression of clock time. Restoration of tissue homeostasis, prevention of degenerative changes, and optimization of collagen alignment hinge on properly dosing mechanical inputs and adjusting rest periods to cellular and molecular demands. The chronobiological perspective suggests that merely counting days or weeks post-injury is insufficient for guiding rehabilitation protocols. Instead, the cumulative number of mechanical load cycles, their magnitude, frequency, and recovery intervals should dictate the progression of therapy [316–318]. Aligning these mechanical cycles with the circadian oscillations governed by BMAL1 and CLOCK may further refine the outcome, making therapies more effective in modulating ECM turnover and cellular metabolism.
Therefore, rehabilitation should not be measured merely by time but by cycles of mechanical loads. By calibrating loading cycles to the ligament’s molecular and cellular rhythms—including circadian gene regulators—clinicians can better facilitate sustainable ECM remodeling, prevent maladaptive tissue responses, and ultimately enhance the durability and functionality of the healing ligament [319–321].
Implications for Tissue Engineering and Regenerative Medicine
The deepening understanding of how ligament cells transduce mechanical signals into molecular and cellular adaptations is providing a robust framework for advanced tissue engineering strategies. In engineered ligament constructs, controlling the mechanical environment at the cellular and molecular levels is key. By using bioreactors that apply physiologically relevant tensile strains to cell-seeded scaffolds, researchers can modulate integrin clustering and focal adhesion formation, guiding cytoskeletal organization and downstream signal transduction [83–85, 322–324]. This mechanical stimulation ensures that cells express the correct repertoire of ECM genes—upregulating collagens I and III, elastin, and proteoglycans—and that newly deposited collagen fibrils become properly aligned, ultimately conferring the scaffold with ligament-like mechanical properties.
Further refinement of scaffold materials can elevate these efforts. Biomaterials that incorporate integrin-binding domains (e.g., RGD motifs) or mimic the native ECM composition provide the essential molecular cues required for proper cell adhesion and mechanosensing [86–88, 325–327]. By presenting ligand-binding sites that selectively engage integrins expressed by ligament fibroblasts, these scaffolds facilitate the formation of focal adhesions and enhance mechanotransduction efficiency. As a result, signaling pathways such as MAPK (ERK, JNK, p38) and PI3K/AKT can be activated more precisely, fine-tuning gene expression and ensuring that the developing construct attains the appropriate density, stiffness, and viscoelastic properties characteristic of native ligaments.
Equally important is the biochemical milieu. Delivering growth factors or cytokines—such as TGF-β, FGF, PDGF, and VEGF—in a controlled manner can further stimulate ECM synthesis, improve collagen fibril organization, and support neovascularization [89–92, 328–330]. Spatially and temporally controlled growth factor release can help synchronize cellular proliferation, differentiation, and matrix remodeling phases, thereby accelerating the ligamentization of tendon grafts or other engineered constructs. Such an approach mirrors in vivo regenerative processes, where growth factor gradients, mechanical load patterns, and cellular signals interact to shape tissue architecture and function [331].
Genetic and epigenetic interventions offer another frontier of possibility for guiding ligament cell fate at the molecular level. For instance, overexpressing Mkx, a transcription factor pivotal for tendon/ligament differentiation, can shift the transcriptome of progenitor cells or tendon-derived cells toward a ligamentous phenotype [93–95, 331–333]. Similarly, modulating microRNA (miRNA) profiles provides a means to fine-tune the balance between anabolic and catabolic processes. By controlling miRNAs that target collagen, MMPs, or growth factor signaling pathways, researchers can encourage stable ECM formation and prevent excessive matrix degradation or scar formation [334–336]. Such genetic or epigenetic “preconditioning” sets the stage for cells to respond optimally once they are placed under mechanical load in a bioreactor or implanted in vivo. With the right molecular switches flipped, the cells become more receptive to integrin-mediated signals, more efficient at organizing ECM, and better able to resist mechanical fatigue. Ultimately, these interventions strive to replicate the intricate molecular milieu and mechanical environment of native ligaments—where integrins, cytoskeletal tension, transcription factors, epigenetic marks, and growth factors interact seamlessly [337–339].
In animals studies the culmination of these efforts is the production of living ligament constructs that not only have the structural and mechanical integrity to replace damaged ligaments but also possess the molecular and cellular machinery to integrate with host tissues and adapt over time [340–342]. However, there is currently no scientific evidence available supporting this approach in humans.
Tendon–Ligament Force Transmission and Weight Saving
Tendons
and ligaments transmit tensile forces across skeletal joints,
effectively redirecting the path of force transmission while allowing
varying degrees of joint motion. Biomechanically, these tissues
feature densely packed, parallel-aligned collagen fibers (primarily
type I collagen), hierarchically organized into microfibrils,
fibrils, and fascicles, interspersed with smaller amounts of elastin
and proteoglycans. This hierarchical organization confers both
strength and flexibility. Ligaments generally function to limit the
range of joint movement and stabilize the joint via their passive
viscoelastic properties [367–369], whereas tendons couple muscle
contraction to bone movement, often spanning one or more joints. From
a molecular standpoint, collagen forms the core of both tendon and
ligament structure. Each type I collagen molecule is composed of
three polypeptide chains (two α1\alpha 1α1 chains and one
α2\alpha 2α2 chain) that form a triple helix. These helices
self-assemble into collagen fibrils, which further aggregate into
collagen fibers. Covalent crosslinks, often catalyzed by lysyl
oxidase, greatly enhance tensile strength and optimize viscoelastic
behavior. The density and nature of these crosslinks help tune creep,
hysteresis, and resilience under cyclical load.
Smaller
molecules such as decorin, biglycan, and fibromodulin (proteoglycans)
help regulate fibril diameter, spacing, and collagen fibrillogenesis.
Meanwhile, glycoproteins (e.g., tenascin-C, fibronectin) mediate cell
adhesion and matrix organization. These constituents also facilitate
water retention and lubrication, contributing to energy dissipation
(damping) and overall tissue viscoelasticity. Fibroblasts (in tendon)
or ligament fibroblasts (sometimes termed “ligamentocytes”)
reside along and within the collagen fascicles. Their integrin
receptors bind ECM molecules, translating mechanical stimuli (e.g.,
stretch, load, shear) into intracellular signals via mechanosensitive
pathways such as focal adhesion kinase (FAK), mitogen-activated
protein kinase (MAPK), and Rho/ROCK signaling. These pathways
orchestrate collagen synthesis, ECM remodeling, and adaptive changes
in matrix composition in response to altered loading regimes.
Such cell–matrix interactions enable tendons and ligaments to undergo homeostatic remodeling, maintaining a balance between matrix deposition and degradation (including matrix metalloproteinase activity). This balance is crucial for sustaining tissue integrity over millions of loading cycles. Under tensile loads, tendons and ligaments can achieve a tensile failure strength of approximately 100–140 MPa and an elastic modulus in the range of 1.0–1.5 GPa [370–373]. This combination of high strength and relatively low mass provides a significant advantage (i.e., “weight savings”) compared to muscle tissue, which is far denser and more metabolically active. Over a given length, one would require roughly 2, 800 times more muscle mass to transmit the same force that a tendon (or ligament) can carry [374, 375]. Consequently, natural selection frequently favors long, slender tendons or ligaments in distal limb segments, especially when fine control is less important than efficient force conduction over distance. Tendon stiffness determines how much the muscle fibers must shorten to produce a given movement. Stiffer tendons transmit force with minimal elongation, whereas more compliant tendons can store and release elastic energy. During activities like running or hopping, this compliance can reduce muscle fiber length changes, thereby decreasing metabolic cost.
By contrast, ligaments generally have slightly lower stiffness and allow controlled elongation, limiting joint excursion while passively contributing to joint stability. They also reduce the need for active muscular stabilization in certain ranges of motion, thus saving metabolic energy.
Extreme examples of the ligamentous strategy appear in the distal limbs of horses and other ungulates (e.g., the digital suspensory ligament), where muscle fibers are vestigial or absent [374, 376–378]. These passive structures transmit force over long distances with minimal mass, significantly lowering energy costs required for posture and locomotion. Humans exhibit a less extreme but still notable reliance on tendons—particularly the Achilles tendon—for storing and releasing elastic energy during gait.
Because tendons and ligaments are metabolically inert in terms of direct ATP consumption, they reduce the overall cost of movement. During gait, tendons can store elastic energy when stretched and release it upon recoil, alleviating the need for muscle fibers to shorten extensively. This mechanism is central to the high efficiency observed in running animals and humans [379–381]. If a tendon is sufficiently compliant, the muscle attached can operate more isometrically, producing force at an optimal fiber length without large length changes. This reduces the metabolic cost of generating force compared to fibers undergoing significant contractions. Over many strides or steps, such energy savings accumulate, enabling greater endurance or speed. A trade-off arises between compliance for energy saving and the capacity for precise control over limb position. Longer, more compliant tendons can introduce delays or oscillations due to the elasticity in the system—particularly relevant when reflex delays (tens of milliseconds) are considered. However, a moderate level of tendon compliance can actually help buffer high-frequency perturbations and refine control of low-force tasks by absorbing sudden shocks [371, 382]. Consequently, muscle–tendon architectures vary across anatomical regions and species, reflecting different functional priorities (e.g., distal vs. proximal limb muscles).
Stress-Strain Curve in Ligaments
Ligaments exhibit a distinctive stress-strain curve that emerges from a sophisticated interplay of molecular components and cellular signaling pathways (Fig. 5). These mechanisms collectively enable ligaments to stabilize joints under various loads while allowing the flexibility required for normal movement [383]. Expanding on traditional views of collagen-dominated mechanics, recent insights reveal that ion channels, mechanosensing molecules, and intricate molecular crosslinking processes are all essential for ligament health and function [384]. The primary load-bearing framework of ligaments is built on collagen, a protein that organizes into a triple-helical structure called tropocollagen [385]. Multiple tropocollagen molecules align into fibrils, which are further grouped into fibers. Ligament collagen is predominantly Type I, conferring high tensile strength, with smaller amounts of Type III contributing to tissue repair and structural support. A hallmark of collagen fibrils is their crimp pattern, a wave-like undulation that is straightened when tension is first applied. During the initial “toe” region of the stress-strain curve, this uncrimping accounts for a notable increase in strain with only a modest rise in stress. As loading continues, collagen fibrils become fully aligned and stiffen the tissue, creating the linear (elastic) region where stress increases sharply with strain [386]. Collagen’s mechanical integrity also relies on covalent crosslinks. Enzymatic crosslinks mediated by lysyl oxidase anchor collagen molecules together, fortifying fibrils against mechanical forces. Over time, non-enzymatic glycation (through advanced glycation end-products, or AGEs (Advanced Glycation End Products) can further increase crosslink density [387]. This extra crosslinking often stiffens ligaments but reduces their extensibility, potentially heightening injury risk and limiting joint mobility, especially in aging tissues or in metabolic conditions such as diabetes [388].
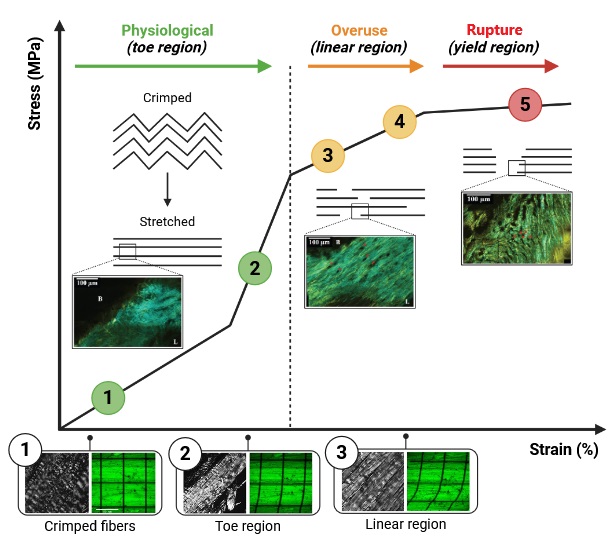
Fig. 5: Depicts the stress-strain curve of ligament tissue, highlighting the progressive molecular and structural changes that occur as ligaments are subjected to varying degrees of mechanical loading. The figure is segmented into five distinct stages, each representing a critical phase in the ligament's response to mechanical stress, ranging from appropriate loading that promotes tissue homeostasis to excessive strain leading to ligament rupture. At the onset, appropriate mechanical loading, characterized by cyclic tensile and shear forces below the threshold for immediate structural failure, activates the PIEZO1 signaling pathway within ligament fibroblasts. PIEZO1, a mechanosensitive ion channel, responds to mechanical stress by facilitating the influx of calcium ions (Ca²⁺) into the cells. This calcium influx triggers downstream signaling cascades involving Calcium/Calmodulin-Dependent Kinases (CaMKs) and Protein Kinase C (PKC), which subsequently activate transcription factors such as CREB and YAP/TAZ. These transcription factors enhance the expression of genes responsible for extracellular matrix (ECM) synthesis, including type I collagen (COL1A1 and COL1A2), elastin, and proteoglycans like decorin and biglycan. The upregulation of these ECM components reinforces the ligament structure, improving its tensile strength and elasticity, thereby maintaining tissue homeostasis and preparing the ligament to better withstand future mechanical loads. As mechanical loading intensifies and strain exceeds the deviation point, the ligament's elastic behavior becomes disrupted. Excessive stress leads to the partial unfolding of collagen triple helices and the disruption of hydrogen bonds that stabilize collagen structures. This molecular unfolding reduces the tensile strength of collagen fibers, while proteoglycans undergo stretching, altering their interactions with collagen and compromising ECM organization. Concurrently, elevated mechanical stress activates the Mitogen-Activated Protein Kinase (MAPK) and Extracellular Signal-Regulated Kinase (ERK) pathways, as well as the NF-κB pathway, which upregulates pro-inflammatory cytokines such as IL-1β and TNF-α. These molecular changes initiate catabolic processes that tilt the balance towards ECM degradation and hinder anabolic repair mechanisms, marking the transition from elastic to plastic deformation of the ligament. Continuing to endure mechanical loading beyond the deviation point, collagen fibers attempt to compensate for the loss of load transfer mechanisms by sliding and stretching at the molecular level. This compensatory behavior generates shear forces that further disrupt the organized alignment of collagen fibrils. The increased shear stress induces the activation of Matrix Metalloproteinases (MMP-2 and MMP-9), which specifically target denatured collagen and gelatin, accelerating ECM degradation. Additionally, mitochondrial dysfunction induced by excessive mechanical stress elevates the production of Reactive Oxygen Species (ROS), contributing to oxidative damage of ECM proteins and cellular components. The combined effect of heightened MMP activity and ROS-mediated damage leads to the gradual degradation of collagen fibers, weakening the structural integrity of fiber bundles and setting the stage for potential rupture. As mechanical loading persists, the gradual degradation of collagen culminates in the rupture of fiber bundles. Sustained high levels of mechanical stress maintain elevated expression of MMP-1 and MMP-13, enzymes that actively degrade type I and type II collagen. Simultaneously, the expression of Tissue Inhibitors of Metalloproteinases (TIMPs) is reduced, diminishing their ability to counteract MMP activity and allowing unchecked ECM degradation. The oxidative environment and pro-inflammatory cytokines activate caspases and regulate BAX/Bcl-2 proteins, leading to programmed cell death (apoptosis) of ligament fibroblasts. The loss of viable fibroblasts impairs the ligament’s capacity to synthesize and maintain ECM components, resulting in the fragmentation of the collagen matrix and the eventual rupture of fiber bundles under continued mechanical strain. Ultimately, accumulated damage from repetitive microstrains below the immediate damage threshold leads to the attainment of a critical damage point. At this juncture, the ligament's ability to remodel and repair the damaged matrix becomes severely compromised. Persistent strain disrupts collagen crosslinks and depletes glycosaminoglycans (GAGs), reducing ECM plasticity and limiting matrix flexibility. The apoptotic loss of fibroblasts and the induction of cellular senescence further diminish ECM synthesis and maintenance, preventing effective repair of microinjuries. Sustained pro-inflammatory cytokine release perpetuates a catabolic environment, exacerbating ECM degradation and inhibiting anabolic processes necessary for ligament healing. Once the critical damage threshold is surpassed, the ligament loses its capacity to undergo necessary plastic deformation, rendering it highly susceptible to sudden rupture even under normal physiological loads. This culmination of molecular and structural degradation underscores the importance of managing mechanical loading to prevent overload-related tendon and ligament disorders. Figure adapter from Eisner LE et al., 2022).
Although collagen is the principal load-bearing component, proteoglycans—comprising a protein core decorated with GAG chains—play critical supporting roles [389]. These negatively charged GAGs attract water and cations, creating a swelling pressure that helps maintain fibril spacing and overall tissue hydration. Water, which makes up the majority of ligament mass, not only lubricates collagen fibrils but also facilitates nutrient transport and dissipates mechanical energy under repeated loading [390]. Glycoproteins like fibronectin and tenascin further organize fibrillar structures and connect cells (primarily fibroblasts) to the extracellular matrix. Although they contribute less to bulk tensile strength than collagen, they help mediate tissue remodeling and repair processes after microinjury [391]. Beyond the structural proteins, ligaments contain resident fibroblasts (sometimes referred to as ligament cells) that continuously monitor and respond to mechanical cues. Mechanosensitive ion channels embedded in fibroblast membranes act as molecular “switches,” transducing mechanical forces into electrochemical signals that guide gene expression and matrix remodeling. Certain TRP subfamily members (e.g., TRPV4) open in response to shear stress or stretch, allowing calcium ions to flow into cells [392]. Increased intracellular calcium can trigger a cascade of signaling events that influence collagen synthesis, proteoglycan turnover, and cytoskeletal reorganization. Piezo1 and Piezo2 are large mechanosensitive ion channels that open upon membrane deformation. When ligaments are subjected to load, stretch-induced Piezo channel activation modulates the release of second messengers, leading to changes in collagen gene expression and cellular proliferation rates. Although not strictly ion channels, integrins—transmembrane receptors linking the extracellular matrix to the cytoskeleton—work in tandem with these channels. As integrins sense tension within collagen fibers, they help coordinate ion channel gating, generating complex feedback loops that ensure tissue homeostasis [393]. Through these ion channel pathways, fibroblasts can sense even small changes in the mechanical environment, adjusting matrix production and degradation to meet physiological demands. Excessive or insufficient mechanical stimulation can shift this balance, potentially weakening the ligament or promoting fibrotic changes [394].
Ligaments are viscoelastic: their response to load varies over time due to fluid movement and molecular rearrangements within the matrix [395]. Water and ions within the proteoglycan-rich network shift as load increases or decreases, allowing ligaments to exhibit phenomena such as creep (gradual elongation under a constant load) and stress relaxation (reduced stress under a constant strain). Collagen molecules can slide and reorient slightly when loaded, enabled by the glycoprotein and proteoglycan interfaces. Over repeated loading cycles, energy is lost (hysteresis) as collagen and matrix components continually adjust. The gating and inactivation rates of mechanosensitive ion channels also contribute to time-dependent behavior [396]. Rapid changes in tension may produce spike-like calcium influxes, while slower deformations can lead to more prolonged changes in channel activity and cell signaling. When load exceeds the ligament’s capacity, plastic (irreversible) deformation and ultimately tissue failure can ensue. Excessive stretching or repetitive loading can break the covalent bonds linking collagen molecules, leading to small-scale microtears. As more crosslinks fail, fibrils lose their aligned structure and the matrix architecture becomes compromised. Activated fibroblasts sense the tissue damage through mechanosensitive channels, potentially upregulating collagen and proteoglycan production in an attempt at repair [397]. However, prolonged or severe overload can overwhelm these responses, resulting in macroscopic tears.
Ligaments do not remain static; they adapt over time and in response to metabolic factors [398]. Aging typically brings heightened collagen crosslink density, reduced proteoglycan content, and more frequent non-enzymatic glycation, causing ligaments to become stiffer and more prone to failure. Diabetes and other metabolic disorders can accelerate glycation, impairing ligament elasticity and contributing to joint dysfunction [399]. Moreover, age- or disease-related changes in ion channel expression can impair fibroblast mechanosensitivity, potentially diminishing the tissue’s capacity to remodel and repair [400]. By recognizing how these molecular features intersect with mechanical load, researchers and clinicians gain critical insights for developing strategies—ranging from pharmacological modulation of ion channels to targeted physical therapies—that preserve ligament integrity and enhance long-term joint function.
Ligament Enthesis: Molecular and Mechanical Transition
The enthesis is a highly specialized anatomical region where ligaments (and tendons) anchor into bone, enabling a seamless transition between soft and hard tissues (Fig. 6). This interface not only confers mechanical resilience by dispersing stress but also embodies a sophisticated molecular and cellular organization essential for tissue homeostasis and repair [401]. Emerging insights from molecular and cellular biology underscore the complexity of enthesis formation and maintenance, with transcription factors (e.g., Scleraxis/Scx, Sox9, Gli-1), growth factors (e.g., GDF-5), and gap junction proteins (Connexins) playing pivotal roles in controlling cell fate, ECM synthesis, and mechanotransduction [402, 403]. Scleraxis is a basic helix-loop-helix transcription factor that is widely regarded as a master regulator of tendon and ligament development. Its expression is predominantly associated with tendon/ligament progenitor cells, where it drives the synthesis of type I collagen and tenascin-C, among other tendon-specific ECM proteins [404]. In the context of the enthesis, Scx-expressing progenitor cells migrate toward the insertion site and initiate the formation of a dense collagen matrix. As mechanical loads intensify, Scx expression can be modulated by integrin-based signaling, ensuring that the enthesis region adapts collagen organization to withstand tensile forces [405].
Fig. 6: Molecular Signaling in the Enthesis and Its Diverse Cell Populations. The enthesis is a biomechanically graded interface where tendons or ligaments merge seamlessly with bone. This transition occurs across distinct zones—fibrous, fibrocartilaginous, and mineralized—that are maintained by specialized cell populations (fibroblast-, chondrocyte-, and osteoblast-like cells, respectively). Numerous molecular signals guide the development, remodeling, and repair of these cell populations, ensuring that the enthesis can withstand multi-axial loading and mitigate stress concentrations. Transcription Factors and Cell Fate Scleraxis (Scx): A master regulator of tendon and ligament development, Scx is vital for synthesizing type I collagen and tendon-specific proteins. Its expression is regulated by mechanical cues via integrin-based signaling, promoting collagen organization in response to tensile forces. Sox9: Primarily associated with chondrogenesis, Sox9 drives the formation of type II collagen and aggrecan, giving fibrocartilage its compressive resistance. Within the enthesis, Sox9 fosters the chondrocyte-like phenotype essential for dampening stress at the transition to mineralized tissue. Gli-1: Functioning downstream of Hedgehog signals, Gli-1-positive progenitors can differentiate into fibroblasts, chondrocytes, or osteoblasts. This dynamic capacity preserves the zonal architecture, allowing ongoing adaptation to mechanical stimuli. Growth Factors and Their Roles GDF-5: A TGF-β superfamily member, GDF-5 enhances progenitor cell recruitment and chondrogenic differentiation. In synergy with BMPs and FGFs, GDF-5 promotes the formation of fibrocartilage, proteoglycan deposition, and eventual mineralization—key for a stable enthesis. Other Signaling Molecules (e.g., TGF-β, BMPs, FGFs): These gradients guide each enthesis zone by modulating anabolic and catabolic processes, harmonizing the production of specialized ECM components. Gap Junction Communication Connexins (Cx43): Gap junctions formed by connexins allow direct exchange of ions and small signaling molecules (e.g., cAMP, IP3) across cells. This intercellular communication is crucial for synchronizing responses to mechanical stress, coordinating ECM turnover, and preventing localized tissue failure. ECM Stratification and Mechanotransduction: ECM Composition: Type I collagen predominates in the tensile ligament region, while type II collagen, type X collagen, and proteoglycans (aggrecan) rise in the fibrocartilage zone, adapting to compressive loads. At the mineralized front, hydroxyapatite integrates with collagen, forming calcified fibrocartilage that transitions to bone. Mechanical Signaling: Integrin-based focal adhesions anchor cells to the ECM and convert mechanical stimuli into biochemical signals. These cues influence transcription factor activity (Scx, Sox9, Gli-1), balancing ECM synthesis and degradation and ensuring the tissue adapts to shifting load demands. Overall, the enthesis’s mechanical resilience relies on finely tuned molecular crosstalk among various cell types. Transcription factors like Scx, Sox9, and Gli-1 orchestrate lineage-specific ECM production, while GDF-5 and other growth factors steer progenitor differentiation. Gap junctions enable rapid signal spread, creating a synchronized response to mechanical stress. Together, these molecular and cellular mechanisms preserve enthesis integrity, enabling fluid load transfer and robust tissue repair when injury occurs.)
Within the fibrocartilaginous region of the enthesis, the transcription factor Sox9 becomes increasingly important. Sox9 is traditionally recognized as a key regulator of chondrogenesis and is essential for the expression of type II collagen and aggrecan [406]. In the transitional zone of the enthesis—where fibrocartilage resides—Sox9 activity helps maintain a phenotype that is partly chondrogenic, providing resistance to compressive forces. This chondrocyte-like phenotype is crucial for reducing stress concentrations at the junction where the ligament transitions into mineralized tissue [407].
Recent studies point to a critical role of Hedgehog signaling in enthesis morphogenesis and maintenance, particularly via the transcription factor Gli-1. Gli-1 is a downstream effector of Sonic hedgehog (Shh) or Indian hedgehog (Ihh) signaling, orchestrating cell proliferation, differentiation, and survival in various skeletal tissues [408]. In the enthesis, Gli-1-positive cells can represent a progenitor population that contributes to ongoing remodeling. These progenitors respond to mechanical cues by modulating their differentiation into fibroblast-, chondrocyte-, or osteoblast-like cells, thus preserving the zonal composition of the ligament-to-bone interface [408, 409]. GDF-5, a member of the transforming growth factor-β (TGF-β) superfamily, has garnered attention for its essential role in joint and tendon/ligament development. In the enthesis, GDF-5 is implicated in stimulating progenitor cell recruitment and chondrogenic differentiation, working synergistically with other factors such as bone morphogenetic proteins (BMPs) and fibroblast growth factors (FGFs) [410]. Experimental models suggest that GDF-5 can promote the formation of fibrocartilaginous zones, enhance proteoglycan deposition, and accelerate the maturation of the mineralized region, all of which are key to a stable enthesis structure [402].
Increasing evidence indicates that enthesis cells rely on gap junction-mediated communication to synchronize matrix synthesis and degradation across different zones. Connexins (particularly Connexin 43, Cx43) form gap junction channels that allow direct intercellular exchange of ions and small molecules, including second messengers like cyclic AMP (cAMP) and inositol triphosphate (IP3) [409]. This connectivity is thought to be crucial for propagating signals that inform cells of mechanical changes, orchestrating a collective response in matrix remodeling. Disruptions in gap junction signaling could lead to maladaptive remodeling and increased susceptibility to enthesopathy or enthesis-related injuries [403]. The ECM at the ligament enthesis is uniquely stratified to handle multiple loading modes. The ligament proper is dominated by type I collagen fibers, which confer tensile strength. Progressing toward the fibrocartilaginous zone, type II collagen, type X collagen, and proteoglycans like aggrecan become more abundant [401, 406]. These components increase the tissue’s capacity to resist compressive forces. At the mineralized interface, hydroxyapatite crystals intercalate with collagen fibers, creating a calcified fibrocartilage region that gradually transitions into subchondral bone [405]. Each of these zones is maintained by a discrete population of cells—fibroblast-like in the ligament region, chondrocyte-like in the fibrocartilage, and osteoblast lineage cells in the bone—which are regulated by molecular gradients involving TGF-β, FGFs, BMPs, and other signaling pathways [407]. The interplay between Scx, Sox9, Gli-1, and GDF-5 ensures that cells in each zone differentiate appropriately and produce ECM components best suited to the mechanical environment [408, 410].
The mechanical environment of the enthesis demands precise modulation of cell behavior. Mechanical loads generated by joint movement are transmitted to enthesis-resident cells, where they are converted into biochemical signals—a process known as mechanotransduction [404]. Integrin-based focal adhesions anchor the cytoskeleton to ECM fibers, enabling cells to sense tension or compression. These signals feed into transcriptional networks involving Scx, Sox9, and Gli-1, adjusting the balance between anabolic (ECM synthesis) and catabolic (ECM breakdown) activities [402].
Further coordination is facilitated by gap junctions composed of Connexins, which allow neighboring cells to share signaling molecules and ions [403]. This rapid intercellular communication ensures that cells within different zones of the enthesis respond cohesively to shifts in mechanical load, thereby preventing localized failure and tissue damage. Finite element analyses and in vivo imaging consistently illustrate that the enthesis is not an abrupt union of two tissues but rather a mechanically graded interface [407]. Such gradation distributes tensile, compressive, and shear forces more evenly across zones, thereby reducing stress concentrations at any single point [401]. The fibrocartilaginous region, in particular, acts as a stress buffer, aided by Sox9- and GDF-5-driven chondrogenic differentiation that boosts proteoglycan content and compressive strength [405, 410].
When mechanical loading patterns change—due to injury or altered joint mechanics—Scx-positive progenitors in the soft tissue region can be activated to reorganize collagen fiber orientation, while Gli-1 and Sox9 pathways adapt the fibrocartilaginous and bone regions to shifting loads [408, 409]. If these adaptive processes are dysregulated (as in enthesitis, osteoarthritis, or chronic overuse), pathological remodeling ensues and may compromise the integrity of the entire insertion site [402].
Future Directions
Future research is poised to significantly expand our molecular and cellular understanding of ligament biology. Single-cell transcriptomics and advanced proteomic profiling will unravel the heterogeneity of ligament cell populations, identifying distinct subpopulations that respond differently to mechanical stimuli [96–98, 343–345]. By examining how individual cells regulate integrin expression, growth factor receptors, and ECM-remodeling enzymes, researchers can pinpoint molecular targets that either enhance ligament resilience or drive degenerative changes. Similarly, high-throughput proteomic analyses can clarify how post-translational modifications (e.g., phosphorylation of FAK or other focal adhesion proteins) and epigenetic regulators (e.g., chromatin-remodeling complexes and miRNAs) shape cell fate decisions under various loading regimens [346–348].
Concurrent advances in imaging technologies will shed light on how these molecular modifications manifest in tissue architecture. Super-resolution microscopy, for instance, can provide nanoscale visualization of integrin clusters, focal adhesions, and cytoskeletal filaments, enabling researchers to link molecular signaling events directly to changes in cell shape and ECM organization [99–101, 349–351]. Second-harmonic generation imaging and other nonlinear optical methods can reveal collagen fiber arrangements in unprecedented detail, correlating molecular-level ECM synthesis and alignment with mechanical properties [352–354]. Such insights can guide the development of scaffolds and rehabilitation programs that precisely modulate cell–ECM interactions.
The integration of computational modeling and machine learning offers a powerful avenue for predicting ligament cell responses to various mechanical loads. By incorporating data from single-cell omics, proteomics, and high-resolution imaging, computational models can simulate how alterations in integrin-binding motifs, growth factor delivery, or microRNA expression affect tissue-level outcomes [102–104, 355–357]. Machine learning algorithms can identify complex patterns in these large datasets, allowing researchers and clinicians to forecast the trajectory of ligament healing, the effectiveness of specific therapeutic interventions, or the optimal mechanical environment for engineered constructs. Tailoring patient-specific rehabilitation protocols and graft-preconditioning strategies becomes feasible as these predictive tools advance, ultimately improving clinical outcomes. As we understand more about how molecular signaling pathways, cellular responses, and mechanical inputs interact, new therapeutic strategies will emerge. Better management of ligament injuries, more efficient ACL graft ligamentization, and refined regenerative medicine techniques will hinge on these molecular insights [358–360]. Clinicians may soon be able to prescribe precision rehabilitation programs that fine-tune mechanical loading parameters based on patient-specific molecular signatures. Similarly, tissue engineers can precondition grafts at the molecular level—overexpressing key transcription factors, adjusting microRNA profiles, or delivering growth factors—to ensure that implanted constructs rapidly adapt to the in vivo environment, producing a functional ligament capable of bearing physiological loads [361–363].
In essence, harnessing these molecular and cellular insights will redefine our approach to ligament repair and regeneration. By integrating cutting-edge omics data, imaging technologies, computational models, and molecular engineering tools, we can move toward a future where ligament injuries are managed with a level of precision and efficacy previously unattainable. This systems-level understanding will bridge the gap between basic mechanobiological research and clinical application, ultimately restoring musculoskeletal function and improving quality of life for patients with ligament injuries [364–366].
Conclusion
This review underscores the complexity and precision with which ligament tissues maintain structural integrity, adapt to their mechanical environment, and respond to injury. At the center of ligament homeostasis lies a delicate interplay between mechanical stimuli, molecular signaling pathways, and cellular phenotypes. Ligament fibroblasts, or ligamentocytes, continuously interpret and translate mechanical load patterns into biochemical responses via integrin-mediated focal adhesions, mechanosensitive ion channels, and associated mechanotransduction networks. The downstream signaling cascades—MAPKs, PI3K/AKT, and others—fine-tune gene expression, guiding collagen fibrillogenesis, ECM composition, and cytoskeletal organization. This robust molecular machinery ensures that ligaments are not passive structures but highly responsive tissues capable of dynamic remodeling.
Distinct from tendons, ligaments must accommodate multidirectional forces. Their unique ECM composition, richer in elastin and proteoglycans, provides compliance and the ability to absorb and dissipate complex stresses. Appropriate mechanical loading preserves collagen alignment and ECM integrity, stabilizing the ligament cell phenotype. Conversely, insufficient or excessive loading triggers inflammatory mediators, catabolic enzymes, and altered transcriptional and epigenetic landscapes, leading to degenerative changes, scar formation, and compromised mechanical properties. Stress shielding and repetitive microtrauma further highlight the importance of balanced mechanical cues. Without the correct level of tension, ligament fibroblasts cannot maintain load-bearing collagen networks, predisposing the tissue to fibrocartilaginous transformation, laxity, or sudden mechanical failure.
Armed with emerging insights into growth factor signaling, cytokine modulation, epigenetic control, and transcriptional regulation, researchers and clinicians can now envision more sophisticated interventions. By modulating integrin ligands, harnessing growth factor gradients, controlling microRNA expression, or applying gene editing tools, it becomes possible to “reprogram” tendon grafts to adopt ligamentous phenotypes, accelerate ACL graft ligamentization, or stabilize degenerative ligaments. Moreover, mechanical stimulation protocols informed by chronobiological principles—calibrating loading cycles rather than relying on static timelines—hold promise for more effective rehabilitation strategies. Such approaches recognize that tissue healing and adaptation depend not solely on the passage of time, but on the cumulative number of load-rest cycles, their intensity, and their coordination with the tissue’s molecular rhythms. These insights directly impact tissue engineering and regenerative medicine. Designing scaffolds equipped with integrin-binding motifs and mechanical cues, delivering controlled growth factors, and employing bioreactors that replicate physiological loading patterns can produce constructs that emulate native ligament properties at the molecular, cellular, and structural levels. By preconditioning cells with genetic or epigenetic interventions, scaffolds can be “tuned” to ensure optimal ECM organization, mechanical resilience, and integration with host tissues post-implantation.
Looking toward the future, advanced single-cell transcriptomics, proteomics, and imaging technologies will deepen our comprehension of ligament cell heterogeneity and molecular dynamics. Combined with computational modeling and machine learning, these data-rich approaches will enable predictive modeling of healing trajectories and patient-specific therapies. Clinicians may soon customize rehabilitation protocols based on molecular “signatures” of a patient’s ligament cells, optimizing load regimens to prevent degenerative cascades and enhance repair. However, it is important to note that there is no single universally optimal mechanical loading regimen; instead, a deep understanding of cellular biology can enhance clinicians’ confidence in selecting targeted rehabilitation approaches and preventing future injuries.
Ultimately, the convergence of molecular mechanobiology, engineering principles, and clinical insight is poised to transform ligament care. However, it is important to note that these concepts remain largely theoretical, as the majority of supporting evidence currently comes from animal models or laboratory-based studies, which may not fully reflect the complexities of human physiology and clinical practice. By bridging fundamental research and clinical application, we can foster a new era of precision medicine in orthopedics—one in which ligament injuries are managed through biologically informed, biomechanically sound strategies that restore function, reduce reinjury risk, and significantly improve patient outcomes.
Abbreviations
ECM (Extracellular Matrix); FAK (Focal Adhesion Kinase); MAPKs (Mitogen-Activated Protein Kinases); PI3K/AKT (Signal Pathways); GAGs (Glycosaminoglycans); SLRPs (Small Leucine-Rich Proteoglycans); TGF-β (Transforming Growth Factor Beta isoforms); FGF (Fibroblast Growth Factor); CTGF (Connective Tissue Growth Factor); MMPs (Matrix Metalloproteinases); TIMPs (Tissue Inhibitors of Metalloproteinases); Scx (Scleraxis Transcription Factor); Mkx (Mohawk Transcription Factor); EGR1 (Early Growth Response 1); GDFs (Growth Differentiation Factors); TRP (Transient Receptor Potential); VEGF (Vascular Endothelial Growth Factor); TFs (Transcription Factors); IGF-1 (Insulin-like Growth Factor 1); NO (Nitric Oxide); ROS ( Reactive Oxygen Species); AGEs (Advanced Glycation End Products)
Disclosure Statement
The authors have no conflicts of interest to declare.
Declaration of AI usage
During
the preparation of this work, the authors used Grammarly to improve
readability and language quality.
References
| 1 | Woo SL, Abramowitch SD, Kilger R, Liang R. Biomechanics of knee ligaments: Injury, healing, and repair. J Biomech 2006;39(1):1-20.
https://doi.org/10.1016/j.jbiomech.2004.10.025 |
| 2 | Benjamin M, Ralphs JR. Fibrocartilage in tendons and ligaments-an adaptation to compressive load. J Anat 1998;193(Pt 4):481-494.
https://doi.org/10.1017/s0021878298004300 |
| 3 | Provenzano PP, Vanderby R Jr. Collagen fibril morphology and organization: Implications for force transmission in ligament and tendon. Matrix Biol 2006;25(2):71-84.
https://doi.org/10.1016/j.matbio.2005.09.005 |
| 4 | Frank CB. Ligament structure, physiology and function. J Musculoskelet Neuronal Interact 2004;4(2):199-201.
|
| 5 | Hoffmann A, Gross G. Tendon and ligament engineering: from cell biology to in vivo application. Regen Med. 2006 Jul;1(4):563-74.
https://doi.org/10.2217/17460751.1.4.563 |
| 6 | Nguyen DT, Dellbrügge S, Tak PP, Woo SL, Blankevoort L, van Dijk NC. Histological characteristics of ligament healing after bio-enhanced repair of the transected goat ACL. J Exp Orthop. 2015 Dec;2(1):4.
https://doi.org/10.1186/s40634-015-0021-5 |
| 7 | Yu X, Hu J, Li Y, Wen Y, Li B. ACL injury management: a comprehensive review of novel biotherapeutics. Front Bioeng Biotechnol. 2024 Nov 22;12:1455225.
https://doi.org/10.3389/fbioe.2024.1455225 |
| 8 | Thorpe CT, Riley GP, Birch HL, Clegg PD, Screen HR. Fascicles and the interfascicular matrix show adaptation for fatigue resistance in energy storing tendons. Acta Biomater 2016;42:308-315.
https://doi.org/10.1016/j.actbio.2016.06.012 |
| 9 | Screen HR. Investigating load relaxation mechanics in tendon. J Mech Behav Biomed Mater 2008;1(1):51-58.
https://doi.org/10.1016/j.jmbbm.2007.03.002 |
| 10 | Kannus P. Structure of the tendon connective tissue. Scand J Med Sci Sports 2000;10(6):312-320.
https://doi.org/10.1034/j.1600-0838.2000.010006312.x |
| 11 | Kjaer M. Role of extracellular matrix in adaptation of tendon and skeletal muscle to mechanical loading. Physiol Rev 2004;84(2):649-698.
https://doi.org/10.1152/physrev.00031.2003 |
| 12 | Thomopoulos S, Marquez JP, Weinberger B, Birman V, Genin GM. Collagen fiber orientation at the tendon to bone insertion. J Biomech 2006;39(10):1842-1851.
https://doi.org/10.1016/j.jbiomech.2005.05.021 |
| 13 | Benjamin M, Kaiser E, Milz S. Structure-function relationships in tendons: A review. J Anat 2008;212(3):211-228.
https://doi.org/10.1111/j.1469-7580.2008.00864.x |
| 14 | Kastelic J, Galeski A, Baer E. The multicomposite structure of tendon. Connect Tissue Res 1978;6(1):11-23.
https://doi.org/10.3109/03008207809152283 |
| 15 | Bramson, M. T. K., Van Houten, S. K., et al. (2021). Mechanobiology in tendon, ligament, and skeletal muscle tissue engineering. Journal of Biomechanical Engineering, 143(7), 070801
https://doi.org/10.1115/1.4050035 |
| 16 | Ingber DE. Cellular mechanotransduction: Putting all the pieces together again. FASEB J 2006;20(7):811-827.
https://doi.org/10.1096/fj.05-5424rev |
| 17 | Franchi, M., Piperigkou, Z., Mastronikolis, N. S., et al. (2024). Extracellular matrix biomechanical roles and adaptation in health and disease. The FEBS Journal
https://doi.org/10.1111/febs.16938 |
| 18 | Wang JH, Thampatty BP. An introductory review of cell mechanobiology. Biomech Model Mechanobiol 2006;5(1):1-16.
https://doi.org/10.1007/s10237-005-0012-z |
| 19 | Geiger B, Spatz JP, Bershadsky AD. Environmental sensing through focal adhesions. Nat Rev Mol Cell Biol 2009;10(1):21-33.
https://doi.org/10.1038/nrm2593 |
| 20 | Li FX, Xu F, Lin X, Wu F, Zhong JY, Wang Y, Guo B, Zheng MH, Shan SK, Yuan LQ. The Role of Substance P in the Regulation of Bone and Cartilage Metabolic Activity. Front Endocrinol (Lausanne). 2020 Feb 28;11:77.
https://doi.org/10.3389/fendo.2020.00077 |
| 21 | Chatterjee, M., Muljadi, P. M., et al. (2022). The role of the tendon ECM in mechanotransduction: Disruption and repair following overuse. Connective Tissue Research, 63(2), 98-110.
https://doi.org/10.1080/03008207.2021.1925663 |
| 22 | Arnoczky SP, Lavagnino M, Egerbacher M. The mechanobiological aetiopathogenesis of tendinopathy: is it the over-stimulation or the under-stimulation of tendon cells? Int J Exp Pathol. 2007 Aug;88(4):217-26.
https://doi.org/10.1111/j.1365-2613.2007.00548.x |
| 23 | Cowin SC. Tissue growth and remodeling. Annu Rev Biomed Eng. 2004;6:77-107.
https://doi.org/10.1146/annurev.bioeng.6.040803.140250 |
| 24 | Burridge K, Guilluy C. Focal adhesions, stress fibers and mechanical tension. Exp Cell Res 2016;343(1):14-20.
https://doi.org/10.1016/j.yexcr.2015.10.029 |
| 25 | Schiller HB, Fässler R. Mechanosensitivity and compositional dynamics of cell-matrix adhesions. EMBO Rep 2013;14(6):509-519.
https://doi.org/10.1038/embor.2013.49 |
| 26 | Parsons JT, Horwitz AR, Schwartz MA. Cell adhesion: Integrating cytoskeletal dynamics and cellular tension. Nat Rev Mol Cell Biol 2010;11(9):633-643.
https://doi.org/10.1038/nrm2957 |
| 27 | Eisner, L. E., Rosario, R., et al. (2022). The role of the non-collagenous extracellular matrix in tendon and ligament mechanical behavior: A review. Journal of Biomechanical Engineering, 144(5), 050801.
https://doi.org/10.1115/1.4053086 |
| 28 | Vogel V, Sheetz M. Local force and geometry sensing regulate cell functions. Nat Rev Mol Cell Biol 2006;7(4):265-275.
https://doi.org/10.1038/nrm1890 |
| 29 | Riveline D, Zamir E, Balaban NQ, et al. Focal contacts as mechanosensors. J Cell Biol 2001;153(5):1175-1185.
https://doi.org/10.1083/jcb.153.6.1175 |
| 30 | Mitra SK, Hanson DA, Schlaepfer DD. Focal adhesion kinase: In command and control of cell motility. Nat Rev Mol Cell Biol 2005;6(1):56-68.
https://doi.org/10.1038/nrm1549 |
| 31 | Eijkelkamp N, Quick K, Wood JN. Transient receptor potential channels and mechanosensation. Annu Rev Neurosci. 2013 Jul 8;36:519-46.
https://doi.org/10.1146/annurev-neuro-062012-170412 |
| 32 | Lambert IH, Hoffmann EK, Pedersen SF. Cell volume regulation: physiology and pathophysiology. Acta Physiol (Oxf). 2008 Dec;194(4):255-82.
https://doi.org/10.1111/j.1748-1716.2008.01910.x |
| 33 | Wu J, Lewis AH, Grandl J. Touch, tension, and transduction - the function and regulation of piezo ion channels. Trends Biochem Sci 2017;42(1):57-71.
https://doi.org/10.1016/j.tibs.2016.09.004 |
| 34 | Praetorius HA, Spring KR. Bending the primary cilium opens Ca²⁺-sensitive intermediate-conductance K⁺ channels in MDCK cells. J Membr Biol 2005;208(2):223-232.
|
| 35 | Delling M, DeCaen PG, Doerner JF, et al. Primary cilia are not calcium-responsive mechanosensors. Nature 2016;531(7596):656-660.
https://doi.org/10.1038/nature17426 |
| 36 | Clark RA. Fibrin and wound healing. Ann N Y Acad Sci 2001;936:355-367.
https://doi.org/10.1111/j.1749-6632.2001.tb03522.x |
| 37 | Sharma P, Maffulli N. Biology of tendon injury: Healing, modeling and remodeling. J Musculoskelet Neuronal Interact 2006;6(2):181-190.
|
| 38 | Chassonnery P, Paupert J, Lorsignol A, Séverac C, Ousset M, Degond P, Casteilla L, Peurichard D. Fibre crosslinking drives the emergence of order in a three-dimensional dynamical network model. R Soc Open Sci. 2024 Jan 31;11(1):231456.
https://doi.org/10.1098/rsos.231456 |
| 39 | Wang JHC, Thampatty BP, Lin JS, Im HJ. Mechanoregulation of gene expression in fibroblasts. Gene 2007;391(1-2):1-15.
https://doi.org/10.1016/j.gene.2007.01.014 |
| 40 | Yang G, Rothrauff BB, Tuan RS. Tendon and ligament regeneration and repair: Clinical relevance and developmental paradigm. Birth Defects Res C Embryo Today 2013;99(3):203-222.
https://doi.org/10.1002/bdrc.21041 |
| 41 | Heinemeier KM, Kjaer M. In vivo investigation of tendon responses to mechanical loading. J Musculoskelet Neuronal Interact 2011;11(2):115-123.
|
| 42 | Plaas AHK, Moran MM, Sandy JD, Hascall VC. Aggrecan and Hyaluronan: The Infamous Cartilage Polyelectrolytes - Then and Now. Adv Exp Med Biol. 2023;1402:3-29.
https://doi.org/10.1007/978-3-031-25588-5_1 |
| 43 | Lavagnino M, Arnoczky SP. In vitro alterations in cytoskeletal tensional homeostasis control gene expression in tendon cells. J Orthop Res 2005;23(5):1211-1218.
https://doi.org/10.1016/j.orthres.2005.04.001 |
| 44 | Killian ML, Thomopoulos S. Scleraxis is required for the development of a functional tendon enthesis. FASEB J. 2016 Jan;30(1):301-11.
https://doi.org/10.1096/fj.14-258236 |
| 45 | Yang F, Zhang A, Richardson DW. Regulation of the tenogenic gene expression in equine tenocyte-derived induced pluripotent stem cells by mechanical loading and Mohawk. Stem Cell Res. 2019 Aug;39:101489.
https://doi.org/10.1016/j.scr.2019.101489 |
| 46 | Tian X, Jiang H, Chen Y, Ao X, Chen C, Zhang W, He F, Liao X, Jiang X, Li T, Zhang Z, Zhang X. Baicalein Accelerates Tendon-Bone Healing via Activation of Wnt/β-Catenin Signaling Pathway in Rats. Biomed Res Int. 2018 Mar 6;2018:3849760.
https://doi.org/10.1155/2018/3849760 |
| 47 | Bi Y, Ehirchiou D, Kilts TM, et al. Identification of tendon stem/progenitor cells and the role of the extracellular matrix niche in their lineage determination. Nat Med 2007;13(10):1219-1227.
https://doi.org/10.1038/nm1630 |
| 48 | Zhang J, Wang JH. The effects of mechanical loading on tendons - an in vivo and in vitro model study. PLoS One 2013;8(8):e71740.
https://doi.org/10.1371/journal.pone.0071740 |
| 49 | Screen HR, Berk DE, Kadler KE, Ramirez F, Young MF. Tendon functional extracellular matrix. J Orthop Res 2015;33(6):793-799.
https://doi.org/10.1002/jor.22818 |
| 50 | Wang T, Chen P, Chen L, Zhou Y, Wang A, Zheng Q, Mitchell CA, Leys T, Tuan RS, Zheng MH. Reduction of mechanical loading in tendons induces heterotopic ossification and activation of the β-catenin signaling pathway. J Orthop Translat. 2021 May 18;29:42-50
https://doi.org/10.1016/j.jot.2021.03.004 |
| 51 | Kjaer M, Heinemeier KM. Eccentric exercise: Acute and chronic effects on healthy and diseased tendons. J Appl Physiol 2014;116(11):1435-1437.
https://doi.org/10.1152/japplphysiol.01044.2013 |
| 52 | Lui PP. Stem cell technology for tendon regeneration. Stem Cells Cloning 2015;8:163-174.
https://doi.org/10.2147/SCCAA.S60832 |
| 53 | Fu SC, Wang W, Pau HM, Wong YP, Chan KM, Rolf CG. Increased expression of transforming growth factor-beta1 in patellar tendinosis. Clin Orthop Relat Res. 2002 Jul;(400):174-83.
https://doi.org/10.1097/00003086-200207000-00022 |
| 54 | Legerlotz K, Riley GP, Screen HR. GAG depletion increases the stress-relaxation response of tendon fascicles, but does not influence recovery. Acta Biomater. 2013 Jun;9(6):6860-6.
https://doi.org/10.1016/j.actbio.2013.02.028 |
| 55 | Millar NL, Akbar M, Campbell AL, Reilly JH, Kerr SC, McLean M, Frleta-Gilchrist M, Fazzi UG, Leach WJ, Rooney BP, Crowe LA, Murrell GA, McInnes IB. IL-17A mediates inflammatory and tissue remodelling events in early human tendinopathy. Sci Rep. 2016 Jun 6;6:27149.
https://doi.org/10.1038/srep27149 |
| 56 | Laurencin CT, Freeman JW. Ligament tissue engineering: An evolutionary materials science approach. Biomaterials 2005;26(36):7530-7536.
https://doi.org/10.1016/j.biomaterials.2005.05.073 |
| 57 | Murray MM, Fleming BC. Use of a collagen-platelet rich plasma scaffold to stimulate healing of a central defect in the ACL. J Orthop Res 2007;25(11):1398-1405.
|
| 58 | Andriacchi TP, Favre J, Erhart-Hledik JC, Chu CR. A systems view of risk factors for knee osteoarthritis reveals insights into the pathogenesis of the disease. Ann Biomed Eng 2015;43(2):376-387.
https://doi.org/10.1007/s10439-014-1117-2 |
| 59 | Luo W, Wang Y, Han Q, Wang Z, Jiao J, Gong X, Liu Y, Zhang A, Zhang H, Chen H, Wang J, Wu M. Advanced strategies for constructing interfacial tissues of bone and tendon/ligament. J Tissue Eng. 2022 Dec 23;13:20417314221144714.
https://doi.org/10.1177/20417314221144714 |
| 60 | Killian ML, Cavinatto L, Galatz LM, Thomopoulos S. The role of mechanobiology in tendon healing and remodeling. J Shoulder Elbow Surg 2012;21(2):228-237.
https://doi.org/10.1016/j.jse.2011.11.002 |
| 61 | Steward RL Jr, Cheng CM, Ye JD, Bellin RM, LeDuc PR. Mechanical stretch and shear flow induced reorganization and recruitment of fibronectin in fibroblasts. Sci Rep. 2011;1:147.
https://doi.org/10.1038/srep00147 |
| 62 | Lui PP, Chan KM. Tendon-derived stem cells (TDSCs): From basic science to potential roles in tendon pathology and tissue engineering applications. Stem Cell Rev Rep 2011;7(4):883-897.
https://doi.org/10.1007/s12015-011-9276-0 |
| 63 | Kuo CK, Tuan RS. Novel strategies in tendon and ligament tissue engineering: Advanced biomaterials and regeneration motifs. Sports Med Arthrosc Rehabil Ther Technol 2010;2:20.
https://doi.org/10.1186/1758-2555-2-20 |
| 64 | Rodeo SA. Biologic augmentation of rotator cuff tendon repair. J Shoulder Elbow Surg 2007;16(5 Suppl):S191-197.
https://doi.org/10.1016/j.jse.2007.03.012 |
| 65 | Connizzo BK, Yannascoli SM, Soslowsky LJ. Structure-function relationships of postnatal tendon development: A parallel to healing. Matrix Biol 2013;32(2):106-116.
https://doi.org/10.1016/j.matbio.2013.01.007 |
| 66 | Boys AJ, McCorry MC, Rodeo S, Bonassar LJ, Estroff LA. Next Generation Tissue Engineering of Orthopedic Soft Tissue-to-Bone Interfaces. MRS Commun. 2017 Sep;7(3):289-308.
https://doi.org/10.1557/mrc.2017.91 |
| 67 | Doroski DM, Brink KS, Temenoff JS. Techniques for biological characterization of tissue-engineered tendon and ligament. Biomaterials. 2007 Jan;28(2):187-202.
https://doi.org/10.1016/j.biomaterials.2006.08.040 |
| 68 | Kjaer M, Magnusson P, Krogsgaard M, Boysen Møller J, Olesen J, Heinemeier K, Hansen M, Haraldsson B, Koskinen S, Esmarck B, Langberg H. Extracellular matrix adaptation of tendon and skeletal muscle to exercise. J Anat. 2006 Apr;208(4):445-50.
https://doi.org/10.1111/j.1469-7580.2006.00549.x |
| 69 | Lavagnino M, Wall ME, Little D, Banes AJ, Guilak F, Arnoczky SP. Tendon mechanobiology: Current knowledge and future research opportunities. J Orthop Res. 2015 Jun;33(6):813-22.
https://doi.org/10.1002/jor.22871 |
| 70 | Benjamin M, Kumai T, Milz S, Boszczyk BM, Boszczyk AA, Ralphs JR. The skeletal attachment of tendons-tendon "entheses." Comp Biochem Physiol A Mol Integr Physiol 2002;133(4):931-945.
https://doi.org/10.1016/S1095-6433(02)00138-1 |
| 71 | Mienaltowski MJ, Gonzales NL, Beall JM, Pechanec MY. Basic Structure, Physiology, and Biochemistry of Connective Tissues and Extracellular Matrix Collagens. Adv Exp Med Biol. 2021;1348:5-43.
https://doi.org/10.1007/978-3-030-80614-9_2 |
| 72 | Golman M, Birman V, Thomopoulos S, Genin GM. Enthesis strength, toughness and stiffness: an image-based model comparing tendon insertions with varying bony attachment geometries. J R Soc Interface. 2021 Dec;18(185):20210421.
https://doi.org/10.1098/rsif.2021.0421 |
| 73 | Leiphart RJ, Pham H, Harvey T, Komori T, Kilts TM, Shetye SS, Weiss SN, Adams SM, Birk DE, Soslowsky LJ, Young MF. Coordinate roles for collagen VI and biglycan in regulating tendon collagen fibril structure and function. Matrix Biol Plus. 2021 Dec 28;13:100099.
https://doi.org/10.1016/j.mbplus.2021.100099 |
| 74 | Smith L, Xia Y, Galatz LM, Genin GM, Thomopoulos S. Tissue-engineering strategies for the tendon/ligament-to-bone insertion. Connect Tissue Res. 2012;53(2):95-105.
https://doi.org/10.3109/03008207.2011.650804 |
| 75 | Wang JH. Mechanobiology of tendon. J Biomech 2006;39(9):1563-1582.
https://doi.org/10.1016/j.jbiomech.2005.05.011 |
| 76 | Liu SH, Yang RS, al-Shaikh R, Lane JM. Collagen in tendon, ligament, and bone healing. A current review. Clin Orthop Relat Res. 1995 Sep;(318):265-78.
|
| 77 | Jung HJ, Fisher MB, Woo SL. Role of biomechanics in the understanding of normal, injured, and healing ligaments and tendons. Sports Med Arthrosc Rehabil Ther Technol. 2009 May 20;1(1):9.
https://doi.org/10.1186/1758-2555-1-9 |
| 78 | Schweitzer R, Chyung JH, Murtaugh LC, et al. Analysis of the tendon cell fate using Scleraxis, a specific marker for tendons and ligaments. Development 2001;128(19):3855-3866.
https://doi.org/10.1242/dev.128.19.3855 |
| 79 | Ito Y, Toriuchi N, Yoshitaka T, et al. The Mohawk homeobox gene is a critical regulator of tendon differentiation. Proc Natl Acad Sci USA 2010;107(23):10538-10542.
https://doi.org/10.1073/pnas.1000525107 |
| 80 | Benhardt HA, Cosgriff-Hernandez EM. The role of mechanical loading in ligament tissue engineering. Tissue Eng Part B Rev. 2009 Dec;15(4):467-75.
https://doi.org/10.1089/ten.teb.2008.0687 |
| 81 | Humphrey JD, Dufresne ER, Schwartz MA. Mechanotransduction and extracellular matrix homeostasis. Nat Rev Mol Cell Biol 2014;15(12):802-812.
https://doi.org/10.1038/nrm3896 |
| 82 | Hynes RO. Integrins: Bidirectional, allosteric signaling machines. Cell 2002;110(6):673-687.
https://doi.org/10.1016/S0092-8674(02)00971-6 |
| 83 | Discher DE, Janmey P, Wang YL. Tissue cells feel and respond to the stiffness of their substrate. Science 2005;310(5751):1139-1143.
https://doi.org/10.1126/science.1116995 |
| 84 | Geiger B, Bershadsky A, Pankov R, Yamada KM. Transmembrane extracellular matrix-cytoskeleton crosstalk. Nat Rev Mol Cell Biol 2001;2(11):793-805.
https://doi.org/10.1038/35099066 |
| 85 | Ingber DE. Tensegrity and mechanotransduction. J Bodyw Mov Ther 2008;12(3):198-200.
https://doi.org/10.1016/j.jbmt.2008.04.038 |
| 86 | Calderwood DA. Integrin activation. J Cell Sci 2004;117(Pt 5):657-666.
https://doi.org/10.1242/jcs.01014 |
| 87 | Mitra SK, Schlaepfer DD. Integrin-regulated FAK-Src signaling in normal and cancer cells. Curr Opin Cell Biol 2006;18(5):516-523.
https://doi.org/10.1016/j.ceb.2006.08.011 |
| 88 | Sieg DJ, Hauck CR, Ilic D, et al. FAK integrates growth-factor and integrin signals to promote cell migration. Nat Cell Biol 2000;2(5):249-256.
https://doi.org/10.1038/35010517 |
| 89 | Parsons JT. Focal adhesion kinase: The first ten years. J Cell Sci 2003;116(Pt 8):1409-1416.
https://doi.org/10.1242/jcs.00373 |
| 90 | Alenghat FJ, Ingber DE. Mechanotransduction: All signals point to cytoskeleton, matrix, and integrins. Sci STKE 2002;2002(119):pe6.
https://doi.org/10.1126/scisignal.1192002pe6 |
| 91 | Azuma N, Duzgun SA, Ikeda M, Kito H, Akasaka N, Sasajima T, Sumpio BE. Endothelial cell response to different mechanical forces. J Vasc Surg. 2000 Oct;32(4):789-94.
https://doi.org/10.1067/mva.2000.107989 |
| 92 | Lee, J.K., Jo, S., Lee, Y.L. et al. Anterior cruciate ligament remnant cells have different potentials for cell differentiation based on their location. Sci Rep 10, 3097 (2020).
https://doi.org/10.1038/s41598-020-60047-w |
| 93 | Ranade SS, Syeda R, Patapoutian A. Mechanically activated ion channels. Neuron 2015;87(6):1162-1179.
https://doi.org/10.1016/j.neuron.2015.08.032 |
| 94 | Coste B, Mathur J, Schmidt M, et al. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010;330(6000):55-60.
https://doi.org/10.1126/science.1193270 |
| 95 | Clapham DE. TRP channels as cellular sensors. Nature 2003;426(6966):517-524.
https://doi.org/10.1038/nature02196 |
| 96 | Lewis AH, Grandl J. Mechanical sensitivity of Piezo1 ion channels can be tuned by cellular membrane tension. Elife 2015;4:e12088.
https://doi.org/10.7554/eLife.12088 |
| 97 | Pathak MM, Nourse JL, Tran T, et al. Stretch-activated ion channel Piezo1 directs lineage choice in human neural stem cells. Proc Natl Acad Sci USA 2014;111(47):16148-16153.
https://doi.org/10.1073/pnas.1409802111 |
| 98 | Martinac B. Mechanosensitive ion channels: Molecules of mechanotransduction. J Cell Sci 2004;117(Pt 12):2449-2460.
https://doi.org/10.1242/jcs.01232 |
| 99 | Ridley AJ. Rho GTPases and actin dynamics in membrane protrusions and vesicle trafficking. Trends Cell Biol 2006;16(10):522-529.
https://doi.org/10.1016/j.tcb.2006.08.006 |
| 100 | Burridge K, Wennerberg K. Rho and Rac take center stage. Cell 2004;116(2):167-179.
https://doi.org/10.1016/S0092-8674(04)00003-0 |
| 101 | Wehrle-Haller B. Assembly and disassembly of cell matrix adhesions. Curr Opin Cell Biol. 2012 Oct;24(5):569-81.
https://doi.org/10.1016/j.ceb.2012.06.010 |
| 102 | Schwartz MA, DeSimone DW. Cell adhesion receptors in mechanotransduction. Curr Opin Cell Biol 2008;20(5):551-556.
https://doi.org/10.1016/j.ceb.2008.05.005 |
| 103 | Bloodgood RA. Sensory reception is an attribute of both primary cilia and motile cilia. J Cell Sci 2010;123(Pt 4):505-509.
https://doi.org/10.1242/jcs.066308 |
| 104 | Satir P, Christensen ST. Overview of structure and function of mammalian cilia. Annu Rev Physiol 2007;69:377-400.
https://doi.org/10.1146/annurev.physiol.69.040705.141236 |
| 105 | Fu S, Lan Y, Wang G, Bao D, Qin B, Zheng Q, Liu H, Wong VKW. External stimulation: A potential therapeutic strategy for tendon-bone healing. Front Bioeng Biotechnol. 2023 Mar 31;11:1150290.
https://doi.org/10.3389/fbioe.2023.1150290 |
| 106 | Bobzin L, Roberts RR, Chen HJ, Crump JG, Merrill AE. Development and maintenance of tendons and ligaments. Development. 2021 Apr 15;148(8):dev186916.
https://doi.org/10.1242/dev.186916 |
| 107 | Lu K, Shi TS, Shen SY, Lu WL, Wu J, Zhang KJ, Zhu XB, Shi Y, Liu XL, Yu F, Li L, Teng HJ, Gao X, Ju HX, Wang W, Li CJ, Jiang Q, Xue B. Egr1 deficiency disrupts dynamic equilibrium of chondrocyte extracellular matrix through PPARγ/RUNX2 signaling pathways. Am J Transl Res. 2018 Jun 15;10(6):1620-1632.
|
| 108 | Dahl KN, Ribeiro AJS, Lammerding J. Nuclear shape, mechanics, and mechanotransduction. Circ Res 2008;102(11):1307-1318.
https://doi.org/10.1161/CIRCRESAHA.108.173989 |
| 109 | Le HQ, Ghatak S, Yeung CY, et al. Mechanical regulation of transcription controls polycomb-mediated gene silencing during lineage commitment. Nat Cell Biol 2016;18(8):864-875.
https://doi.org/10.1038/ncb3387 |
| 110 | Millar NL, Gilchrist DS, Akbar M, Reilly JH, Kerr SC, Campbell AL, Murrell GAC, Liew FY, Kurowska-Stolarska M, McInnes IB. MicroRNA29a regulates IL-33-mediated tissue remodelling in tendon disease. Nat Commun. 2015 Apr 10;6:6774.
https://doi.org/10.1038/ncomms7774 |
| 111 | Son YH, Yang DH, Uricoli B, Park SJ, Jeong GJ, Chun HJ. Three-Dimensional Cell Culture System for Tendon Tissue Engineering. Tissue Eng Regen Med. 2023 Jul;20(4):553-562.
https://doi.org/10.1007/s13770-023-00550-z |
| 112 | Trumbull A, Subramanian G, Yildirim-Ayan E. Mechanoresponsive musculoskeletal tissue differentiation of adipose-derived stem cells. Biomed Eng Online. 2016 Apr 22;15:43.
https://doi.org/10.1186/s12938-016-0150-9 |
| 113 | Siadat, S.M., Zamboulis, D.E., Thorpe, C.T., Ruberti, J.W., Connizzo, B.K. (2021). Tendon Extracellular Matrix Assembly, Maintenance and Dysregulation Throughout Life. In: Halper, J. (eds) Progress in Heritable Soft Connective Tissue Diseases. Advances in Experimental Medicine and Biology, vol 1348. Springer, Cham.
https://doi.org/10.1007/978-3-030-80614-9_3 |
| 114 | Banos CC, Thomas AH, Kuo CK. Collagen fibrillogenesis in tendon development: current models and regulation of fibril assembly. Birth Defects Res C Embryo Today. 2008 Sep;84(3):228-44.
https://doi.org/10.1002/bdrc.20130 |
| 115 | Debnath B, Narasimhan BN, Fraley SI, Rangamani P. Modeling collagen fibril degradation as a function of matrix microarchitecture. bioRxiv [Preprint]. 2024 Aug 12:2024.08.10.607470.
https://doi.org/10.1101/2024.08.10.607470 |
| 116 | Kongsgaard M, Aagaard P, Kjaer M, Magnusson SP. Structural Achilles tendon properties in athletes subjected to different exercise modes and in Achilles tendon rupture patients. J Appl Physiol 2005;99(5):1965-1971.
https://doi.org/10.1152/japplphysiol.00384.2005 |
| 117 | Lu HH, Thomopoulos S. Functional attachment of soft tissues to bone: development, healing, and tissue engineering. Annu Rev Biomed Eng. 2013;15:201-26.
https://doi.org/10.1146/annurev-bioeng-071910-124656 |
| 118 | Delgado Caceres M, Pfeifer CG, Docheva D. Understanding Tendons: Lessons from Transgenic Mouse Models. Stem Cells Dev. 2018 Sep 1;27(17):1161-1174.
https://doi.org/10.1089/scd.2018.0121 |
| 119 | Gaut L, Robert N, Delalande A, Bonnin MA, Pichon C, Duprez D. EGR1 Regulates Transcription Downstream of Mechanical Signals during Tendon Formation and Healing. PLoS One. 2016 Nov 7;11(11):e0166237.
https://doi.org/10.1371/journal.pone.0166237 |
| 120 | Pintucci G, Steinberg BM, Seghezzi G, Yun J, Apazidis A, Baumann FG, Grossi EA, Colvin SB, Mignatti P, Galloway AC. Mechanical endothelial damage results in basic fibroblast growth factor-mediated activation of extracellular signal-regulated kinases. Surgery. 1999 Aug;126(2):422-7.
https://doi.org/10.1016/S0039-6060(99)70187-X |
| 121 | Ogata Y, Mabuchi Y, Shinoda K, Horiike Y, Mizuno M, Otabe K, Suto EG, Suzuki N, Sekiya I, Akazawa C. Anterior cruciate ligament-derived mesenchymal stromal cells have a propensity to differentiate into the ligament lineage. Regen Ther. 2018 Jan 23;8:20-28.
https://doi.org/10.1016/j.reth.2017.12.001 |
| 122 | Wang JH, Thampatty BP. Mechanobiology of adult and stem cells. Int Rev Cell Mol Biol 2008;271:301-346.
https://doi.org/10.1016/S1937-6448(08)01207-0 |
| 123 | Rod E, Matić I, Antunović M, Vetma V, Pavičić I, Hudetz D, Marijanović I, Primorac D, Ivković A. Optimization of an ex vivo gene transfer to the hamstrings tendons muscle remnants: potential for genetic enhancement of bone healing. Croat Med J. 2019 Jun 13;60(3):201-211.
https://doi.org/10.3325/cmj.2019.60.201 |
| 124 | Juneja SC, Veillette C. Defects in tendon, ligament, and enthesis in response to genetic alterations in key proteoglycans and glycoproteins: a review. Arthritis. 2013;2013:154812.
https://doi.org/10.1155/2013/154812 |
| 125 | Walia B, Li TM, Crosio G, Montero AM, Huang AH. Axin2-lineage cells contribute to neonatal tendon regeneration. Connect Tissue Res. 2022 Sep;63(5):530-543.
https://doi.org/10.1080/03008207.2022.2036732 |
| 126 | Li Y, Yao L, Huang Y, Pang L, Zhang C, Li T, Wang D, Zhou K, Li J, Tang X. Leptin Enhances M1 Macrophage Polarization and Impairs Tendon-Bone Healing in Rotator Cuff Repair: A Rat Model. Clin Orthop Relat Res. 2025 Feb 19;483(5):939-51.
https://doi.org/10.1097/CORR.0000000000003428 |
| 127 | Canty EG, Lu Y, Meadows RS, Shaw MK, Holmes DF, Kadler KE. Coalignment of plasma membrane channels and protrusions (fibripositors) specifies the parallelism of tendon. J Cell Biol. 2004 May 24;165(4):553-63
https://doi.org/10.1083/jcb.200312071 |
| 128 | Riley GP. Tendinopathy-from basic science to treatment. Nat Clin Pract Rheumatol 2008;4(2):82-89.
https://doi.org/10.1038/ncprheum0700 |
| 129 | Thorpe CT, Birch HL, Clegg PD, Screen HR. The role of the non-collagenous matrix in tendon function. Int J Exp Pathol 2013;94(4):248-259.
https://doi.org/10.1111/iep.12027 |
| 130 | Millar NL, Hueber AJ, Reilly JH, Xu Y, Murrell GA, McInnes IB. Inflammation is present in early human tendinopathy. Am J Sports Med 2010;38(10):2085-2091.
https://doi.org/10.1177/0363546510372613 |
| 131 | Tang C, Chen Y, Huang J, Zhao K, Chen X, Yin Z, Heng BC, Chen W, Shen W. The roles of inflammatory mediators and immunocytes in tendinopathy. J Orthop Translat. 2018 Apr 14;14:23-33.
https://doi.org/10.1016/j.jot.2018.03.003 |
| 132 | Hamilton B, Purdam C. Patellar tendinosis as an adaptive process: a new hypothesis. Br J Sports Med. 2004 Dec;38(6):758-61.
https://doi.org/10.1136/bjsm.2003.005157 |
| 133 | de Mos M, Joosten LA, Oppers-Walgreen B, van Schie JT, Jahr H, van Osch GJ, Verhaar JA. Tendon degeneration is not mediated by regulation of Toll-like receptors 2 and 4 in human tenocytes. J Orthop Res. 2009 Aug;27(8):1043-7.
https://doi.org/10.1002/jor.20834 |
| 134 | Lyu K, Liu X, Liu T, Lu J, Jiang L, Chen Y, Long L, Wang X, Shi H, Wang F, Li S. miRNAs contributing to the repair of tendon injury. Cell Tissue Res. 2023 Aug;393(2):201-215.
https://doi.org/10.1007/s00441-023-03780-8 |
| 135 | Cuevas-Cervera M, Aguilar-Nuñez D, Aguilar-García M, García-Ríos MC, González-Muñoz A, Navarro-Ledesma S. Patellar Tendon Elasticity and Temperature Following after a 448 Kilohertz Radiofrequency Intervention on Active Healthy Subjects: An Open Controlled Clinical Trial. Diagnostics (Basel). 2023 Sep 18;13(18):2976.
https://doi.org/10.3390/diagnostics13182976 |
| 136 | Woo SL, Hollis JM, Adams DJ, Lyon RM, Takai S. Tensile properties of the human femur-anterior cruciate ligament-tibia complex: The effects of specimen age and orientation. Am J Sports Med 1991;19(3):217-225.
https://doi.org/10.1177/036354659101900303 |
| 137 | Amiel D, Frank CB, Harwood FL, et al. Tendons and ligaments: A morphological and biochemical comparison. J Orthop Res 1984;1(3):257-265.
https://doi.org/10.1002/jor.1100010305 |
| 138 | Heinemeier KM, Schjerling P, Heinemeier J, Magnusson SP, Kjaer M. Lack of tissue renewal in human adult Achilles tendon is revealed by nuclear bomb (14)C. FASEB J. 2013 May;27(5):2074-9.
https://doi.org/10.1096/fj.12-225599 |
| 139 | Claes S, Verdonk P, Forsyth R, Bellemans J. The "Ligamentization" process in ACL reconstruction: What happens to the human graft? A systematic review of the literature. Am J Sports Med 2011;39(11):2476-2483.
https://doi.org/10.1177/0363546511402662 |
| 140 | Moretti L, Bizzoca D, Cassano GD, Caringella N, Delmedico M, Moretti B. Graft Intra-Articular Remodeling and Bone Incorporation in ACL Reconstruction: The State of the Art and Clinical Implications. Journal of Clinical Medicine. 2022; 11(22):6704.
https://doi.org/10.3390/jcm11226704 |
| 141 | Kanazawa T, Soejima T, Murakami H, Inoue T, Katouda M, Nagata K. An immunohistological study of the integration at the bone-tendon interface after reconstruction of the anterior cruciate ligament in rabbits. J Bone Joint Surg Br. 2006;88(5):682-7.
https://doi.org/10.1302/0301-620X.88B5.17198 |
| 142 | Goradia VK, Rochat MC, Grana WA, Rohrer MD, Prasad HS. Tendon-to-bone healing of a semitendinosus tendon autograft used for ACL reconstruction in a sheep model. Am J Knee Surg. 2000 Summer;13(3):143-51.
|
| 143 | Holzapfel GA, Humphrey JD, Ogden RW. Biomechanics of soft biological tissues and organs, mechanobiology, homeostasis and modelling. J R Soc Interface. 2025 Jan;22(222):20240361.
https://doi.org/10.1098/rsif.2024.0361 |
| 144 | Chiquet M, Renedo AS, Huber F, Flück M. How do fibroblasts translate mechanical signals into changes in extracellular matrix production? Matrix Biol 2003;22(1):73-80.
https://doi.org/10.1016/S0945-053X(03)00004-0 |
| 145 | Anderson T, Mo J, Gagarin E, Sherwood D, Blumenkrantz M, Mao E, Leon G, Levitz H, Chen HJ, Tseng KC, Fabian P, Crump JG, Smeeton J. Ligament injury in adult zebrafish triggers ECM remodeling and cell dedifferentiation for scar-free regeneration. NPJ Regen Med. 2023 Sep 19;8(1):51.
https://doi.org/10.1038/s41536-023-00329-9 |
| 146 | Wu YF, Wang HK, Chang HW, Sun J, Sun JS, Chao YH. High glucose alters tendon homeostasis through downregulation of the AMPK/Egr1 pathway. Sci Rep. 2017 Mar 7;7:44199.
https://doi.org/10.1038/srep44199 |
| 147 | Pentzold S, Wildemann B. Mechanical overload decreases tenogenic differentiation compared to physiological load in bioartificial tendons. J Biol Eng. 2022 Mar 3;16(1):5.
https://doi.org/10.1186/s13036-022-00283-y |
| 148 | Muhammad H, Rais Y, Miosge N, Ornan EM. The primary cilium as a dual sensor of mechanochemical signals in chondrocytes. Cell Mol Life Sci. 2012 Jul;69(13):2101-7.
https://doi.org/10.1007/s00018-011-0911-3 |
| 149 | Wang T, Wang L, Zhang L, Long Y, Zhang Y, Hou Z. Single-cell RNA sequencing in orthopedic research. Bone Res. 2023 Feb 24;11(1):10.
https://doi.org/10.1038/s41413-023-00245-0 |
| 150 | Koob TJ, Summers AP. Tendon--bridging the gap. Comp Biochem Physiol A Mol Integr Physiol. 2002 Dec;133(4):905-9.
https://doi.org/10.1016/S1095-6433(02)00255-6 |
| 151 | Galloway MT, Lalley AL, Shearn JT. The role of mechanical loading in tendon development, maintenance, injury, and repair. J Bone Joint Surg Am. 2013 Sep 4;95(17):1620-8.
https://doi.org/10.2106/JBJS.L.01004 |
| 152 | Evans CH, Huard J. Gene therapy approaches to regenerating the musculoskeletal system. Nat Rev Rheumatol. 2015 Apr;11(4):234-42.
https://doi.org/10.1038/nrrheum.2015.28 |
| 153 | Iozzo RV. Matrix proteoglycans: From molecular design to cellular function. Annu Rev Biochem 1998;67:609-652.
https://doi.org/10.1146/annurev.biochem.67.1.609 |
| 154 | Thomopoulos S, Genin GM, Galatz LM. The development and morphogenesis of the tendon-to-bone insertion - what development can teach us about healing -. J Musculoskelet Neuronal Interact. 2010 Mar;10(1):35-45.
|
| 155 | Liu Y, Ramanath HS, Wang DA. Tendon tissue engineering using scaffold enhancing strategies. Trends Biotechnol 2008;26(4):201-209.
https://doi.org/10.1016/j.tibtech.2008.01.003 |
| 156 | Kjaer M, Langberg H, Heinemeier K, et al. From mechanical loading to collagen synthesis, structural changes and function in human tendon. Scand J Med Sci Sports 2009;19(4):500-510.
https://doi.org/10.1111/j.1600-0838.2009.00986.x |
| 157 | Aureille J, Belaadi N, Guilluy C. Mechanotransduction via the nuclear envelope: a distant reflection of the cell surface. Curr Opin Cell Biol. 2017 Feb;44:59-67.
https://doi.org/10.1016/j.ceb.2016.10.003 |
| 158 | Guo Y, Calve S, Tepole AB. Multiscale mechanobiology: Coupling models of adhesion kinetics and nonlinear tissue mechanics. Biophys J. 2022 Feb 15;121(4):525-539.
https://doi.org/10.1016/j.bpj.2022.01.012 |
| 159 | Millar NL, Gilchrist DS, Akbar M, Reilly JH, Kerr SC, Campbell AL, Murrell GAC, Liew FY, Kurowska-Stolarska M, McInnes IB. MicroRNA29a regulates IL-33-mediated tissue remodelling in tendon disease. Nat Commun. 2015 Apr 10;6:6774.
https://doi.org/10.1038/ncomms7774 |
| 160 | Paxton JZ, Hagerty P, Andrick JJ, Baar K. Optimizing an intermittent stretch paradigm using ERK1/2 phosphorylation results in increased collagen synthesis in engineered ligaments. Tissue Eng Part A. 2012 Feb;18(3-4):277-84.
https://doi.org/10.1089/ten.tea.2011.0336 |
| 161 | Vunjak-Novakovic G, Altman G, Horan R, Kaplan DL. Tissue engineering of ligaments. Annu Rev Biomed Eng. 2004;6:131-56.
https://doi.org/10.1146/annurev.bioeng.6.040803.140037 |
| 162 | Sahoo S, Toh SL, Goh JC. A bFGF-releasing silk/PLGA-based biohybrid scaffold for ligament/tendon tissue engineering using mesenchymal progenitor cells. Biomaterials 2010;31(11):2990-2998.
https://doi.org/10.1016/j.biomaterials.2010.01.004 |
| 163 | Perucca Orfei C, Bowles AC, Kouroupis D, Willman MA, Ragni E, Kaplan LD, Best TM, Correa D, de Girolamo L. Human Tendon Stem/Progenitor Cell Features and Functionality Are Highly Influenced by in vitro Culture Conditions. Front Bioeng Biotechnol. 2021 Sep 20;9:711964.
https://doi.org/10.3389/fbioe.2021.711964 |
| 164 | Sopilidis A, Stamatopoulos V, Giannatos V, Taraviras G, Panagopoulos A, Taraviras S. Integrating Modern Technologies into Traditional Anterior Cruciate Ligament Tissue Engineering. Bioengineering. 2025; 12(1):39.
https://doi.org/10.3390/bioengineering12010039 |
| 165 | Shi BY, Sriram V, Wu SY, Huang D, Cheney A, Metzger MF, Sundberg O, Lyons KM, McKenna CE, Nishimura I, Kremen TJ Jr. Novel bisphosphonate-based cathepsin K-triggered compound targets the enthesis without impairing soft tissue-to-bone healing. Front Bioeng Biotechnol. 2024 Feb 16;12:1308161.
https://doi.org/10.3389/fbioe.2024.1308161 |
| 166 | Citro V, Clerici M, Boccaccini AR, Della Porta G, Maffulli N, Forsyth NR. Tendon tissue engineering: An overview of biologics to promote tendon healing and repair. J Tissue Eng. 2023 Sep 13;14:20417314231196275.
https://doi.org/10.1177/20417314231196275 |
| 167 | Asahara H, Inui M, Lotz MK. Tendons and Ligaments: Connecting Developmental Biology to Musculoskeletal Disease Pathogenesis. J Bone Miner Res. 2017 Sep;32(9):1773-1782.
https://doi.org/10.1002/jbmr.3199 |
| 168 | Felsenthal N, Zelzer E. Mechanical regulation of musculoskeletal system development. Development. 2017 Dec 1;144(23):4271-4283.
https://doi.org/10.1242/dev.151266 |
| 169 | Nagakura R, Yamamoto M, Jeong J, Hinata N, Katori Y, Chang WJ, Abe S. Switching of Sox9 expression during musculoskeletal system development. Sci Rep. 2020 May 21;10(1):8425.
https://doi.org/10.1038/s41598-020-65339-9 |
| 170 | Lee KI, Gamini R, Olmer M, Ikuta Y, Hasei J, Baek J, Alvarez-Garcia O, Grogan SP, D'Lima DD, Asahara H, Su AI, Lotz MK. Mohawk is a transcription factor that promotes meniscus cell phenotype and tissue repair and reduces osteoarthritis severity. Sci Transl Med. 2020 Oct 28;12(567):eaan7967.
https://doi.org/10.1126/scitranslmed.aan7967 |
| 171 | Havis E, Duprez D. EGR1 Transcription Factor is a Multifaceted Regulator of Matrix Production in Tendons and Other Connective Tissues. Int J Mol Sci. 2020 Feb 28;21(5):1664.
https://doi.org/10.3390/ijms21051664 |
| 172 | Kshitiz, Kim DH, Beebe DJ, Levchenko A. Micro- and nanoengineering for stem cell biology: the promise with a caution. Trends Biotechnol. 2011 Aug;29(8):399-408.
https://doi.org/10.1016/j.tibtech.2011.03.006 |
| 173 | Umur E, Bulut SB, Yiğit P, Bayrak E, Arkan Y, Arslan F, Baysoy E, Kaleli-Can G, Ayan B. Exploring the Role of Hormones and Cytokines in Osteoporosis Development. Biomedicines. 2024 Aug 12;12(8):1830.
https://doi.org/10.3390/biomedicines12081830 |
| 174 | Dupont S, Morsut L, Aragona M, et al. Role of YAP/TAZ in mechanotransduction. Nature 2011;474(7350):179-183.
https://doi.org/10.1038/nature10137 |
| 175 | Panciera T, Azzolin L, Cordenonsi M, Piccolo S. Mechanobiology of YAP and TAZ in physiology and disease. Nat Rev Mol Cell Biol 2017;18(12):758-770.
https://doi.org/10.1038/nrm.2017.87 |
| 176 | Janota CS, Calero-Cuenca FJ, Gomes ER. The role of the cell nucleus in mechanotransduction. Curr Opin Cell Biol. 2020 Apr;63:204-211.
https://doi.org/10.1016/j.ceb.2020.03.001 |
| 177 | Xu Y, Huang Y, Cheng X, Hu B, Jiang D, Wu L, Peng S, Hu J. Mechanotransductive receptor Piezo1 as a promising target in the treatment of fibrosis diseases. Front Mol Biosci. 2023 Oct 12;10:1270979.
https://doi.org/10.3389/fmolb.2023.1270979 |
| 178 | Miroshnikova YA, Nava MM, Wickström SA. Emerging roles of mechanical forces in chromatin regulation. J Cell Sci 2017;130(14):2243-2250.
https://doi.org/10.1242/jcs.202192 |
| 179 | Uhler C, Shivashankar GV. Regulation of genome organization and gene expression by nuclear mechanotransduction. Nat Rev Mol Cell Biol 2017;18(12):717-727.
https://doi.org/10.1038/nrm.2017.101 |
| 180 | Iyer KV, Pulford S, Mogilner A, Shivashankar GV. Mechanical activation of cells induces chromatin remodeling preceding MKL nuclear transport. Biophys J 2012;103(7):1416-1428.
https://doi.org/10.1016/j.bpj.2012.08.041 |
| 181 | Zhang J, Nie D, Williamson K, McDowell A, Hogan MV, Wang JH. Moderate and intensive mechanical loading differentially modulate the phenotype of tendon stem/progenitor cells in vivo. PLoS One. 2020 Dec 29;15(12):e0242640.
https://doi.org/10.1371/journal.pone.0242640 |
| 182 | Liu Q, Zhu Y, Zhu W, Zhang G, Yang YP, Zhao C. The role of MicroRNAs in tendon injury, repair, and related tissue engineering. Biomaterials. 2021 Oct;277:121083.
https://doi.org/10.1016/j.biomaterials.2021.121083 |
| 183 | Chainani PH, Buzo Mena M, Yeritsyan D, Caro D, Momenzadeh K, Galloway JL, DeAngelis JP, Ramappa AJ, Nazarian A. Successive tendon injury in an in vivo rat overload model induces early damage and acute healing responses. Front Bioeng Biotechnol. 2024 Mar 7;12:1327094.
https://doi.org/10.3389/fbioe.2024.1327094 |
| 184 | Brinlee AW, Dickenson SB, Hunter-Giordano A, Snyder-Mackler L. ACL Reconstruction Rehabilitation: Clinical Data, Biologic Healing, and Criterion-Based Milestones to Inform a Return-to-Sport Guideline. Sports Health. 2022 Sep-Oct;14(5):770-779.
https://doi.org/10.1177/19417381211056873 |
| 185 | Yang Y, Wu Y, Zhou K, Wu D, Yao X, Heng BC, Zhou J, Liu H, Ouyang H. Interplay of Forces and the Immune Response for Functional Tendon Regeneration. Front Cell Dev Biol. 2021 Jun 4;9:657621.
https://doi.org/10.3389/fcell.2021.657621 |
| 186 | Donderwinkel I, Tuan RS, Cameron NR, Frith JE. Tendon tissue engineering: Current progress towards an optimized tenogenic differentiation protocol for human stem cells. Acta Biomater. 2022 Jun;145:25-42.
https://doi.org/10.1016/j.actbio.2022.04.028 |
| 187 | Liu X, Shiwaku Y, Hamai R, Tsuchiya K, Takahashi T, Suzuki O. Effect of Octacalcium Phosphate Crystals on the Osteogenic Differentiation of Tendon Stem/Progenitor Cells In Vitro. Int J Mol Sci. 2023 Jan 8;24(2):1235.
https://doi.org/10.3390/ijms24021235 |
| 188 | Schulze-Tanzil GG. Healing of Ligaments and Tendons: Tissue Engineering and Models. Int J Mol Sci. 2022 Dec 7;23(24):15503.
https://doi.org/10.3390/ijms232415503 |
| 189 | Ingber DE, Folkman J. Mechanochemical switching between growth and differentiation during fibroblast growth factor-stimulated angiogenesis in vitro: role of extracellular matrix. J Cell Biol. 1989 Jul;109(1):317-30.
https://doi.org/10.1083/jcb.109.1.317 |
| 190 | De R, Zemel A, Safran SA. Theoretical concepts and models of cellular mechanosensing. Methods Cell Biol. 2010;98:143-75.
https://doi.org/10.1016/S0091-679X(10)98007-2 |
| 191 | Wan Q, Kim SJ, Yokota H, Na S. Differential activation and inhibition of RhoA by fluid flow induced shear stress in chondrocytes. Cell Biol Int. 2013 Jun;37(6):568-76.
https://doi.org/10.1002/cbin.10072 |
| 192 | Sivaramakrishnan S, Spudich JA. Systematic control of protein interaction using a modular ER/K α-helix linker. Proc Natl Acad Sci U S A. 2011 Dec 20;108(51):20467-72.
https://doi.org/10.1073/pnas.1116066108 |
| 193 | Heyn JCJ, Rädler JO, Falcke M. Mesenchymal cell migration on one-dimensional micropatterns. Front Cell Dev Biol. 2024 Apr 16;12:1352279.
https://doi.org/10.3389/fcell.2024.1352279 |
| 194 | Chen CS, Tan J, Tien J. Mechanotransduction at cell-matrix and cell-cell contacts. Annu Rev Biomed Eng 2004;6:275-302.
https://doi.org/10.1146/annurev.bioeng.6.040803.140040 |
| 195 | Watanabe T, Kametani K, Koyama YI, Suzuki D, Imamura Y, Takehana K, Hiramatsu K. Ring-Mesh Model of Proteoglycan Glycosaminoglycan Chains in Tendon based on Three-dimensional Reconstruction by Focused Ion Beam Scanning Electron Microscopy. J Biol Chem. 2016 Nov 4;291(45):23704-23708.
https://doi.org/10.1074/jbc.M116.733857 |
| 196 | Chen Z, Jin M, He H, Dong J, Li J, Nie J, Wang Z, Xu J, Wu F. Mesenchymal stem cells and macrophages and their interactions in tendon-bone healing. J Orthop Translat. 2023 Jan 20;39:63-73
https://doi.org/10.1016/j.jot.2022.12.005 |
| 197 | Gögele C, Hoffmann C, Konrad J, Merkel R, Schwarz S, Tohidnezhad M, Hoffmann B, Schulze-Tanzil GG. Cyclically stretched ACL fibroblasts emigrating from spheroids adapt their cytoskeleton and ligament-related expression profile. Cell Tissue Res. 2021 Jun;384(3):675-690.
https://doi.org/10.1007/s00441-021-03416-9 |
| 198 | Ornitz DM, Itoh N. Fibroblast growth factors. Genome Biol 2001;2(3):REVIEWS3005.
https://doi.org/10.1186/gb-2001-2-3-reviews3005 |
| 199 | Subramanian A, Schilling TF. Tendon development and musculoskeletal assembly: emerging roles for the extracellular matrix. Development. 2015 Dec 15;142(24):4191-204.
https://doi.org/10.1242/dev.114777 |
| 200 | Font Tellado S, Chiera S, Bonani W, Poh PSP, Migliaresi C, Motta A, Balmayor ER, van Griensven M. Heparin functionalization increases retention of TGF-β2 and GDF5 on biphasic silk fibroin scaffolds for tendon/ligament-to-bone tissue engineering. Acta Biomater. 2018 May;72:150-166.
https://doi.org/10.1016/j.actbio.2018.03.017 |
| 201 | Adam NC, Smith CR, Herzog W, Amis AA, Arampatzis A, Taylor WR. In Vivo Strain Patterns in the Achilles Tendon During Dynamic Activities: A Comprehensive Survey of the Literature. Sports Med Open. 2023 Jul 19;9(1):60.
https://doi.org/10.1186/s40798-023-00604-5 |
| 202 | Calvo-Rubio M, Garcia-Domiguez E, Tamayo-Torres E, Soto-Rodríguez S, Olaso-Gonzalez G, Ferrucci L, de Cabo R, Gómez-Cabrera MC. The repeated bout effect evokes the training-induced skeletal muscle cellular memory. Free Radic Biol Med. 2024 Nov 20;225:247-254.
https://doi.org/10.1016/j.freeradbiomed.2024.09.047 |
| 203 | Doessing S, Heinemeier KM, Holm L, Mackey AL, Schjerling P, Rennie M, Smith K, Reitelseder S, Kappelgaard AM, Rasmussen MH, Flyvbjerg A, Kjaer M. Growth hormone stimulates the collagen synthesis in human tendon and skeletal muscle without affecting myofibrillar protein synthesis. J Physiol. 2010 Jan 15;588(Pt 2):341-51.
https://doi.org/10.1113/jphysiol.2009.179325 |
| 204 | Roseti L, Buda R, Cavallo C, Desando G, Facchini A, Grigolo B. Ligament repair: a molecular and immunohistological characterization. J Biomed Mater Res A. 2008 Jan;84(1):117-27.
https://doi.org/10.1002/jbm.a.31449 |
| 205 | Cook JL, Rio E, Purdam CR, Docking SI. Revisiting the continuum model of tendon pathology: what is its merit in clinical practice and research? Br J Sports Med. 2016 Oct;50(19):1187-91.
https://doi.org/10.1136/bjsports-2015-095422 |
| 206 | Guerquin MJ, Charvet B, Nourissat G, Havis E, Ronsin O, Bonnin MA, Ruggiu M, Olivera-Martinez I, Robert N, Lu Y, Kadler KE, Baumberger T, Doursounian L, Berenbaum F, Duprez D. Transcription factor EGR1 directs tendon differentiation and promotes tendon repair. J Clin Invest. 2013 Aug;123(8):3564-76.
https://doi.org/10.1172/JCI67521 |
| 207 | Shahid H, Morya VK, Oh JU, Kim JH, Noh KC. Hypoxia-Inducible Factor and Oxidative Stress in Tendon Degeneration: A Molecular Perspective. Antioxidants (Basel). 2024 Jan 10;13(1):86.
https://doi.org/10.3390/antiox13010086 |
| 208 | Wang Z, Ma C, Chen D, Haslett C, Xu C, Dong C, Wang X, Zheng M, Jing Y, Feng JQ. Tendon Cells Root Into (Instead of Attach to) Humeral Bone Head via Fibrocartilage-Enthesis. Int J Biol Sci. 2023 Jan 1;19(1):183-203.
https://doi.org/10.7150/ijbs.79007 |
| 209 | Wang H, He K, Cheng CK. The Structure, Biology, and Mechanical Function of Tendon/Ligament-Bone Interfaces. Tissue Eng Part B Rev. 2024 Oct;30(5):545-558.
https://doi.org/10.1089/ten.teb.2023.0295 |
| 210 | Roemer FW, Englund M, Turkiewicz A, Struglics A, Guermazi A, Lohmander LS, Larsson S, Frobell R. Molecular and Structural Biomarkers of Inflammation at Two Years After Acute Anterior Cruciate Ligament Injury Do Not Predict Structural Knee Osteoarthritis at Five Years. Arthritis Rheumatol. 2019 Feb;71(2):238-243.
https://doi.org/10.1002/art.40687 |
| 211 | Shepherd JH, Legerlotz K, Demirci T, Klemt C, Riley GP, Screen HR. Functionally distinct tendon fascicles exhibit different creep and stress relaxation behaviour. Proc Inst Mech Eng H. 2014 Jan;228(1):49-59.
https://doi.org/10.1177/0954411913509977 |
| 212 | Scott A, Lian Ø, Roberts CR, Cook JL, Handley CJ, Bahr R, Samiric T, Ilic MZ, Parkinson J, Hart DA, Duronio V, Khan KM. Increased versican content is associated with tendinosis pathology in the patellar tendon of athletes with jumper's knee. Scand J Med Sci Sports. 2008 Aug;18(4):427-35.
https://doi.org/10.1111/j.1600-0838.2007.00735.x |
| 213 | Zhu YX, Huang JQ, Ming YY, Zhuang Z, Xia H. Screening of key biomarkers of tendinopathy based on bioinformatics and machine learning algorithms. PLoS One. 2021 Oct 29;16(10):e0259475.
https://doi.org/10.1371/journal.pone.0259475 |
| 214 | Giordano L, Porta GD, Peretti GM, Maffulli N. Therapeutic potential of microRNA in tendon injuries. Br Med Bull. 2020 May 15;133(1):79-94.
https://doi.org/10.1093/bmb/ldaa002 |
| 215 | Aggouras AN, Stowe EJ, Mlawer SJ, Connizzo BK. Aged Tendons Exhibit Altered Mechanisms of Strain-Dependent Extracellular Matrix Remodeling. J Biomech Eng. 2024 Jul 1;146(7):071009.
https://doi.org/10.1115/1.4065270 |
| 216 | Lin X, Ayrapetov MK, Lee S, Parang K, Sun G. Probing the communication between the regulatory and catalytic domains of a protein tyrosine kinase, Csk. Biochemistry. 2005 Feb 8;44(5):1561-7.
https://doi.org/10.1021/bi048142j |
| 217 | Immanuel CN, Teng B, Dong B, Gordon EM, Kennedy JA, Luellen C, Schwingshackl A, Cormier SA, Fitzpatrick EA, Waters CM. Apoptosis signal-regulating kinase-1 promotes inflammasome priming in macrophages. Am J Physiol Lung Cell Mol Physiol. 2019 Mar 1;316(3):L418-L427.
https://doi.org/10.1152/ajplung.00199.2018 |
| 218 | Clancy RM, Abramson SB. Nitric oxide: a novel mediator of inflammation. Proc Soc Exp Biol Med. 1995 Nov;210(2):93-101.
https://doi.org/10.3181/00379727-210-43927AA |
| 219 | Kim SE, Kim JG, Park K. Biomaterials for the Treatment of Tendon Injury. Tissue Eng Regen Med. 2019 Sep 13;16(5):467-477.
https://doi.org/10.1007/s13770-019-00217-8 |
| 220 | Sugita S, Suzumura T, Nakamura A, Tsukiji S, Ujihara Y, Nakamura M. Second harmonic generation light quantifies the ratio of type III to total (I + III) collagen in a bundle of collagen fiber. Sci Rep. 2021 Jun 4;11(1):11874.
https://doi.org/10.1038/s41598-021-91302-3 |
| 221 | Weidlich K, Mersmann F, Domroes T, Schroll A, Bohm S, Arampatzis A. Quantification of patellar tendon strain and opportunities for personalized tendon loading during back squats. Sci Rep. 2023 May 29;13(1):8661
https://doi.org/10.1038/s41598-023-35441-9 |
| 222 | Osmanagic-Myers S, Dechat T, Foisner R. Lamins at the crossroads of mechanosignaling. Genes Dev. 2015 Feb 1;29(3):225-37.
https://doi.org/10.1101/gad.255968.114 |
| 223 | Yeh CF, Chou C, Yang KC. Mechanotransduction in fibrosis: Mechanisms and treatment targets. Curr Top Membr. 2021;87:279-314.
https://doi.org/10.1016/bs.ctm.2021.07.004 |
| 224 | Gögele C, Vogt J, Hahn J, Breier A, Bernhardt R, Meyer M, Schröpfer M, Schäfer-Eckart K, Schulze-Tanzil G. Co-Culture of Mesenchymal Stem Cells and Ligamentocytes on Triphasic Embroidered Poly(L-lactide-co-ε-caprolactone) and Polylactic Acid Scaffolds for Anterior Cruciate Ligament Enthesis Tissue Engineering. Int J Mol Sci. 2023 Apr 4;24(7):6714.
https://doi.org/10.3390/ijms24076714 |
| 225 | Grier WG, Moy AS, Harley BA. Cyclic tensile strain enhances human mesenchymal stem cell Smad 2/3 activation and tenogenic differentiation in anisotropic collagen-glycosaminoglycan scaffolds. Eur Cell Mater. 2017 Mar 20;33:227-239.
https://doi.org/10.22203/eCM.v033a17 |
| 226 | Full SM, Delman C, Gluck JM, Abdmaulen R, Shemin RJ, Heydarkhan-Hagvall S. Effect of fiber orientation of collagen-based electrospun meshes on human fibroblasts for ligament tissue engineering applications. J Biomed Mater Res B Appl Biomater. 2015 Jan;103(1):39-46.
https://doi.org/10.1002/jbm.b.33153 |
| 227 | Ho JO, Sawadkar P, Mudera V. A review on the use of cell therapy in the treatment of tendon disease and injuries. J Tissue Eng. 2014 Sep 18;5:2041731414549678.
https://doi.org/10.1177/2041731414549678 |
| 228 | Wang J, Xu J, Fu W, Cheng W, Chan K, Yung PS, Qin L. Biodegradable Magnesium Screws Accelerate Fibrous Tissue Mineralization at the Tendon-Bone Insertion in Anterior Cruciate Ligament Reconstruction Model of Rabbit. Sci Rep. 2017 Jan 10;7:40369.
https://doi.org/10.1038/srep40369 |
| 229 | Kaji DA, Montero AM, Patel R, Huang AH. Transcriptional profiling of mESC-derived tendon and fibrocartilage cell fate switch. Nat Commun. 2021 Jul 9;12(1):4208.
https://doi.org/10.1038/s41467-021-24535-5 |
| 230 | Hee CK, Dines JS, Dines DM, Roden CM, Wisner-Lynch LA, Turner AS, McGilvray KC, Lyons AS, Puttlitz CM, Santoni BG. Augmentation of a rotator cuff suture repair using rhPDGF-BB and a type I bovine collagen matrix in an ovine model. Am J Sports Med. 2011 Aug;39(8):1630-9.
https://doi.org/10.1177/0363546511404942 |
| 231 | Lu J, Chen H, Lyu K, Jiang L, Chen Y, Long L, Wang X, Shi H, Li S. The Functions and Mechanisms of Tendon Stem/Progenitor Cells in Tendon Healing. Stem Cells Int. 2023 Sep 12;2023:1258024.
https://doi.org/10.1155/2023/1258024 |
| 232 | Yasuda K, Hayashi K. Changes in biomechanical properties of tendons and ligaments from joint disuse. Osteoarthritis Cartilage. 1999 Jan;7(1):122-9.
https://doi.org/10.1053/joca.1998.0167 |
| 233 | Kanazawa T, Nakagami G, Minematsu T, Yamane T, Huang L, Mugita Y, Noguchi H, Mori T, Sanada H. Biological responses of three-dimensional cultured fibroblasts by sustained compressive loading include apoptosis and survival activity. PLoS One. 2014 Aug 7;9(8):e104676.
https://doi.org/10.1371/journal.pone.0104676 |
| 234 | Hwang J, Lee SY, Jo CH. Degenerative tendon matrix induces tenogenic differentiation of mesenchymal stem cells. J Exp Orthop. 2023 Feb 14;10(1):15.
https://doi.org/10.1186/s40634-023-00581-4 |
| 235 | Bell JS, Hayes S, Whitford C, Sanchez-Weatherby J, Shebanova O, Terrill NJ, Sørensen TLM, Elsheikh A, Meek KM. Tropocollagen springs allow collagen fibrils to stretch elastically. Acta Biomater. 2022 Apr 1;142:185-193.
https://doi.org/10.1016/j.actbio.2022.01.041 |
| 236 | Ekdahl M, Wang JH, Ronga M, Fu FH. Graft healing in anterior cruciate ligament reconstruction. Knee Surg Sports Traumatol Arthrosc. 2008 Oct;16(10):935-47.
https://doi.org/10.1007/s00167-008-0584-0 |
| 237 | Filbay SR, Grindem H. Evidence-based recommendations for the management of anterior cruciate ligament (ACL) rupture. Best Pract Res Clin Rheumatol. 2019 Feb;33(1):33-47.
https://doi.org/10.1016/j.berh.2019.01.018 |
| 238 | Fu W, Li Q, Tang X, Chen G, Zhang C, Li J. Mesenchymal stem cells reside in anterior cruciate ligament remnants in situ. Int Orthop. 2016 Jul;40(7):1523-30.
https://doi.org/10.1007/s00264-015-2925-1 |
| 239 | Chen Z, Chen P, Zheng M, Gao J, Liu D, Wang A, Zheng Q, Leys T, Tai A, Zheng M. Challenges and perspectives of tendon-derived cell therapy for tendinopathy: from bench to bedside. Stem Cell Res Ther. 2022 Sep 2;13(1):444.
https://doi.org/10.1186/s13287-022-03113-6 |
| 240 | Russo V, El Khatib M, Prencipe G, Citeroni MR, Faydaver M, Mauro A, Berardinelli P, Cerveró-Varona A, Haidar-Montes AA, Turriani M, Di Giacinto O, Raspa M, Scavizzi F, Bonaventura F, Stöckl J, Barboni B. Tendon Immune Regeneration: Insights on the Synergetic Role of Stem and Immune Cells during Tendon Regeneration. Cells. 2022 Jan 27;11(3):434.
https://doi.org/10.3390/cells11030434 |
| 241 | Liu CF, Breidenbach A, Aschbacher-Smith L, Butler D, Wylie C. A role for hedgehog signaling in the differentiation of the insertion site of the patellar tendon in the mouse. PLoS One. 2013 Jun 10;8(6):e65411.
https://doi.org/10.1371/journal.pone.0065411 |
| 242 | da Silva MR, Andrade R, Cardoso FS, Oliveira S, Catarino SO, Carvalho Ó, Silva FS, Espregueira-Mendes J, Flores P. Light stimulation on tenocytes: A systematic review of in vitro studies. Porto Biomed J. 2022 Sep 9;7(4):e176.
https://doi.org/10.1097/j.pbj.0000000000000176 |
| 243 | Abraham AC, Shah SA, Golman M, Song L, Li X, Kurtaliaj I, Akbar M, Millar NL, Abu-Amer Y, Galatz LM, Thomopoulos S. Targeting the NF-κB signaling pathway in chronic tendon disease. Sci Transl Med. 2019 Feb 27;11(481):eaav4319.
https://doi.org/10.1126/scitranslmed.aav4319 |
| 244 | Fang F, Sup M, Luzzi A, Ferrer X, Thomopoulos S. Hedgehog signaling underlying tendon and enthesis development and pathology. Matrix Biol. 2022 Jan;105:87-103.
https://doi.org/10.1016/j.matbio.2021.12.001 |
| 245 | Majima T, Funakoshi T, Suenaga N, et al. Alginate and chitosan polyion complex hybrid fibers for scaffolds in ligament and tendon tissue engineering. J Orthop Sci 2005;10(3):302-307.
https://doi.org/10.1007/s00776-005-0891-y |
| 246 | Grier WK, Sun Han Chang RA, Ramsey MD, Harley BAC. The influence of cyclic tensile strain on multi-compartment collagen-GAG scaffolds for tendon-bone junction repair. Connect Tissue Res. 2019 Nov;60(6):530-543.
https://doi.org/10.1080/03008207.2019.1601183 |
| 247 | Jacques P, McGonagle D. The role of mechanical stress in the pathogenesis of spondyloarthritis and how to combat it. Best Pract Res Clin Rheumatol. 2014 Oct;28(5):703-10.
https://doi.org/10.1016/j.berh.2014.10.009 |
| 248 | Vervaecke AJ, Carbone AD, Abraham A, Bernstein Z, Laudier D, Verborgt O, Galatz LM, Huang AH. Tendon progenitor cells as biological augmentation improve functional gait and reduce scar formation after rotator cuff repair. J Shoulder Elbow Surg. 2022 Nov;31(11):2366-2380.
https://doi.org/10.1016/j.jse.2022.05.004 |
| 249 | Puetzer JL, Ma T, Sallent I, Gelmi A, Stevens MM. Driving Hierarchical Collagen Fiber Formation for Functional Tendon, Ligament, and Meniscus Replacement. Biomaterials. 2021 Feb;269:120527.
https://doi.org/10.1016/j.biomaterials.2020.120527 |
| 250 | Hannafin JA, Arnoczky SP, Hooimeyer J, Torzilli PA, Warren RF. Effect of stress deprivation and cyclic tensile loading on the material and mechanical properties of canine flexor digitorum profundus tendons. J Orthop Res 1995;13(6):907-914.
https://doi.org/10.1002/jor.1100130615 |
| 251 | Ingber DE. Tensegrity-based mechanosensing from macro to micro. Prog Biophys Mol Biol 2008;97(2-3):163-179.
https://doi.org/10.1016/j.pbiomolbio.2008.02.005 |
| 252 | Guilak F, Cohen DM, Estes BT, Gimble JM, Liedtke W, Chen CS. Control of stem cell fate by physical interactions with the extracellular matrix. Cell Stem Cell. 2009 Jul 2;5(1):17-26.
https://doi.org/10.1016/j.stem.2009.06.016 |
| 253 | Sotodosos-Alonso L, Pulgarín-Alfaro M, Del Pozo MA. Caveolae Mechanotransduction at the Interface between Cytoskeleton and Extracellular Matrix. Cells. 2023 Mar 20;12(6):942.
https://doi.org/10.3390/cells12060942 |
| 254 | Atta G, Schroedl F, Kaser-Eichberger A, Spitzer G, Traweger A, Heindl LM, Tempfer H. Scleraxis expressing scleral cells respond to inflammatory stimulation. Histochem Cell Biol. 2021 Aug;156(2):123-132.
https://doi.org/10.1007/s00418-021-01985-y |
| 255 | Wang JH, Li B. Mechanics rules cell biology. Sports Med Arthrosc Rehabil Ther Technol. 2010 Jul 8;2:16. doi: 10.1186/1758-2555-2-16.
https://doi.org/10.1186/1758-2555-2-16 |
| 256 | Gross G, Hoffmann A. Therapeutic strategies for tendon healing based on novel biomaterials, factors and cells. Pathobiology. 2013;80(4):203-10.
https://doi.org/10.1159/000347059 |
| 257 | Kjaer M, Langberg H, Miller BF, Boushel R, Crameri R, Koskinen S, Heinemeier K, Olesen JL, Døssing S, Hansen M, Pedersen SG, Rennie MJ, Magnusson P. Metabolic activity and collagen turnover in human tendon in response to physical activity. J Musculoskelet Neuronal Interact. 2005 Mar;5(1):41-52.
|
| 258 | Kondratko-Mittnacht J, Lakes R, Vanderby R Jr. Shear loads induce cellular damage in tendon fascicles. J Biomech. 2015 Sep 18;48(12):3299-305.
https://doi.org/10.1016/j.jbiomech.2015.06.006 |
| 259 | Liu Y, Schwartz AG, Hong Y, Peng X, Xu F, Thomopoulos S, Genin GM. Correction of bias in the estimation of cell volume fraction from histology sections. J Biomech. 2020 May 7;104:109705.
https://doi.org/10.1016/j.jbiomech.2020.109705 |
| 260 | Yu CH, Luo W, Sheetz MP. Spatial-temporal reorganization of activated integrins. Cell Adh Migr. 2012 May-Jun;6(3):280-4.
https://doi.org/10.4161/cam.20753 |
| 261 | Schmitt MS, Colen J, Sala S, Devany J, Seetharaman S, Caillier A, Gardel ML, Oakes PW, Vitelli V. Machine learning interpretable models of cell mechanics from protein images. Cell. 2024 Jan 18;187(2):481-494.e24.
https://doi.org/10.1016/j.cell.2023.11.041 |
| 262 | Chen CS. Mechanotransduction-a field pulling together? J Cell Sci 2008;121(Pt 20):3285-3292.
https://doi.org/10.1242/jcs.023507 |
| 263 | Siadat SM, Zamboulis DE, Thorpe CT, Ruberti JW, Connizzo BK. Tendon Extracellular Matrix Assembly, Maintenance and Dysregulation Throughout Life. Adv Exp Med Biol. 2021;1348:45-103.
https://doi.org/10.1007/978-3-030-80614-9_3 |
| 264 | Arnoczky SP, Lavagnino M, Egerbacher M, Caballero O, Gardner K, Shender MA. Loss of homeostatic strain alters mechanostat "set point" of tendon cells in vitro. Clin Orthop Relat Res. 2008 Jul;466(7):1583-91.
https://doi.org/10.1007/s11999-008-0264-x |
| 265 | Wang HN, Huang YC, Ni GX. Mechanotransduction of stem cells for tendon repair. World J Stem Cells. 2020 Sep 26;12(9):952-965.
https://doi.org/10.4252/wjsc.v12.i9.952 |
| 266 | De Corato M, Gomez-Benito MJ. Interplay of chromatin organization and mechanics of the cell nucleus. Biophys J. 2024 Oct 1;123(19):3386-3396.
https://doi.org/10.1016/j.bpj.2024.08.003 |
| 267 | Davidson PM, Lammerding J. Broken nuclei--lamins, nuclear mechanics, and disease. Trends Cell Biol. 2014 Apr;24(4):247-56.
https://doi.org/10.1016/j.tcb.2013.11.004 |
| 268 | Wen D, Gao Y, Ho C, Yu L, Zhang Y, Lyu G, Hu D, Li Q, Zhang Y. Focusing on Mechanoregulation Axis in Fibrosis: Sensing, Transduction and Effecting. Front Mol Biosci. 2022 Mar 11;9:804680.
https://doi.org/10.3389/fmolb.2022.804680 |
| 269 | Coste B. Détecter la pression? Identification de deux protéines activées par les forces mécaniques [Feeling the pressure? Identification of two proteins activated by mechanical forces]. Med Sci (Paris). 2011 Jan;27(1):17-9. French.
https://doi.org/10.1051/medsci/201127117 |
| 270 | Huang J, Korsunsky A, Yazdani M, Chen J. Targeting TRP channels: recent advances in structure, ligand binding, and molecular mechanisms. Front Mol Neurosci. 2024 Jan 11;16:1334370.
https://doi.org/10.3389/fnmol.2023.1334370 |
| 271 | Khalilimeybodi A, Fraley SI, Rangamani P. Mechanisms underlying divergent relationships between Ca2+ and YAP/TAZ signalling. J Physiol. 2023 Feb;601(3):483-515. doi: 10.1113/JP283966.
https://doi.org/10.1113/JP283966 |
| 272 | Guo J, Tang H, Huang P, Ye X, Tang C, Shu Z, Guo J, Kang X, Shi Y, Zhou B, Liang T, Tang K. Integrative single-cell RNA and ATAC sequencing reveals that the FOXO1-PRDX2-TNF axis regulates tendinopathy. Front Immunol. 2023 May 8;14:1092778.
https://doi.org/10.3389/fimmu.2023.1092778 |
| 273 | Abdalla AA, Pendegrass CJ. Biological approaches to the repair and regeneration of the rotator cuff tendon-bone enthesis: a literature review. Biomater Transl. 2023 Jun 28;4(2):85-103.
|
| 274 | Susilo ME, Paten JA, Sander EA, Nguyen TD, Ruberti JW. Collagen network strengthening following cyclic tensile loading. Interface Focus. 2016 Feb 6;6(1):20150088.
https://doi.org/10.1098/rsfs.2015.0088 |
| 275 | Lazarczuk SL, Maniar N, Opar DA, Duhig SJ, Shield A, Barrett RS, Bourne MN. Mechanical, Material and Morphological Adaptations of Healthy Lower Limb Tendons to Mechanical Loading: A Systematic Review and Meta-Analysis. Sports Med. 2022 Oct;52(10):2405-2429.
https://doi.org/10.1007/s40279-022-01695-y |
| 276 | Grodman LH, Beaulieu ML, Ashton-Miller JA, Wojtys EM. Levels of ACL-straining activities increased in the six months prior to non-contact ACL injury in a retrospective survey: evidence consistent with ACL fatigue failure. Front Physiol. 2023 May 5;14:1166980.
https://doi.org/10.3389/fphys.2023.1166980 |
| 277 | Data K, Kulus M, Ziemak H, Chwarzyński M, Piotrowska-Kempisty H, Bukowska D, Antosik P, Mozdziak P, Kempisty B. Decellularization of Dense Regular Connective Tissue-Cellular and Molecular Modification with Applications in Regenerative Medicine. Cells. 2023 Sep 16;12(18):2293.
https://doi.org/10.3390/cells12182293 |
| 278 | Wudebwe UN, Bannerman A, Goldberg-Oppenheimer P, Paxton JZ, Williams RL, Grover LM. Exploiting cell-mediated contraction and adhesion to structure tissues in vitro. Philos Trans R Soc Lond B Biol Sci. 2015 Feb 5;370(1661):20140200.
https://doi.org/10.1098/rstb.2014.0200 |
| 279 | Bernotiene E, Bagdonas E, Kirdaite G, Bernotas P, Kalvaityte U, Uzieliene I, Thudium CS, Hannula H, Lorite GS, Dvir-Ginzberg M, Guermazi A, Mobasheri A. Emerging Technologies and Platforms for the Immunodetection of Multiple Biochemical Markers in Osteoarthritis Research and Therapy. Front Med (Lausanne). 2020 Oct 21;7:572977.
https://doi.org/10.3389/fmed.2020.572977 |
| 280 | Kular JK, Basu S, Sharma RI. The extracellular matrix: Structure, composition, age-related differences, tools for analysis and applications for tissue engineering. J Tissue Eng. 2014 Dec 20;5:2041731414557112.
https://doi.org/10.1177/2041731414557112 |
| 281 | Lin TW, Cardenas L, Soslowsky LJ. Biomechanics of tendon injury and repair. J Biomech. 2004 Jun;37(6):865-77.
https://doi.org/10.1016/j.jbiomech.2003.11.005 |
| 282 | Mengsteab PY, Conroy P, Badon M, Otsuka T, Kan HM, Vella AT, Nair LS, Laurencin CT. Evaluation of a bioengineered ACL matrix's osteointegration with BMP-2 supplementation. PLoS One. 2020 Jan 7;15(1):e0227181.
https://doi.org/10.1371/journal.pone.0227181 |
| 283 | Ingber DE. Cellular tensegrity: Defining new rules of biological design that govern the cytoskeleton. J Cell Sci 1993;104(Pt 3):613-627.
https://doi.org/10.1242/jcs.104.3.613 |
| 284 | Smeets JSJ, Horstman AMH, Vles GF, Emans PJ, Goessens JPB, Gijsen AP, van Kranenburg JMX, van Loon LJC. Protein synthesis rates of muscle, tendon, ligament, cartilage, and bone tissue in vivo in humans. PLoS One. 2019 Nov 7;14(11):e0224745.
https://doi.org/10.1371/journal.pone.0224745 |
| 285 | Leahy TP, Chenna SS, Soslowsky LJ, Dyment NA. Focal adhesion kinase regulates tendon cell mechanoresponse and physiological tendon development. FASEB J. 2024 Sep;38(17):e70050.
https://doi.org/10.1096/fj.202400151R |
| 286 | Young KA, Biggins L, Sharpe HJ. Protein tyrosine phosphatases in cell adhesion. Biochem J. 2021 Mar 12;478(5):1061-1083.
https://doi.org/10.1042/BCJ20200511 |
| 287 | Zhu L, Plow EF, Qin J. Initiation of focal adhesion assembly by talin and kindlin: A dynamic view. Protein Sci. 2021 Mar;30(3):531-542.
https://doi.org/10.1002/pro.4014 |
| 288 | Kolasangiani R, Bidone TC, Schwartz MA. Integrin Conformational Dynamics and Mechanotransduction. Cells. 2022 Nov 12;11(22):3584.
https://doi.org/10.3390/cells11223584 |
| 289 | Du Y, Xu B, Li Q, Peng C, Yang K. The role of mechanically sensitive ion channel Piezo1 in bone remodeling. Front Bioeng Biotechnol. 2024 Feb 8;12:1342149.
https://doi.org/10.3389/fbioe.2024.1342149 |
| 290 | McEntire DM, Kirkpatrick DR, Dueck NP, Kerfeld MJ, Smith TA, Nelson TJ, Reisbig MD, Agrawal DK. Pain transduction: a pharmacologic perspective. Expert Rev Clin Pharmacol. 2016 Aug;9(8):1069-80.
https://doi.org/10.1080/17512433.2016.1183481 |
| 291 | Zhang M, Meng N, Wang X, Chen W, Zhang Q. TRPV4 and PIEZO Channels Mediate the Mechanosensing of Chondrocytes to the Biomechanical Microenvironment. Membranes (Basel). 2022 Feb 18;12(2):237.
https://doi.org/10.3390/membranes12020237 |
| 292 | Thankam FG, Dilisio MF, Gross RM, Agrawal DK. Collagen I: a kingpin for rotator cuff tendon pathology. Am J Transl Res. 2018 Nov 15;10(11):3291-3309.
|
| 293 | Yang G, Lin H, Rothrauff BB, Yu S, Tuan RS. Multilayered polycaprolactone/gelatin fiber-hydrogel composite for tendon tissue engineering. Acta Biomater. 2016 Apr 15;35:68-76.
https://doi.org/10.1016/j.actbio.2016.03.004 |
| 294 | Han SH, Kim HK, Ahn JH, Lee DH, Baek M, Ye G, Lee JM, Min K, Oh C, Lee S. A Protocol to Acquire the Degenerative Tenocyte from Humans. J Vis Exp. 2018 Jun 9;(136):57634.
https://doi.org/10.3791/57634-v |
| 295 | Locke RC, Abraham AC, Killian ML. Orthopedic Interface Repair Strategies Based on Native Structural and Mechanical Features of the Multiscale Enthesis. ACS Biomater Sci Eng. 2017 Nov 13;3(11):2633-2643.
https://doi.org/10.1021/acsbiomaterials.6b00599 |
| 296 | Collins DW, Diaz MA, Coutelle N, McLaughlin M, Pamic D, Simon P, Doarn MC. Biomechanical Evaluation of Novel Tendon Coaptation Technique: A Potential Application for Tendon Transfers. J Hand Surg Glob Online. 2024 Dec 10;7(2):152-157.
https://doi.org/10.1016/j.jhsg.2024.11.007 |
| 297 | Backman LJ, Eriksson DE, Danielson P. Substance P reduces TNF-α-induced apoptosis in human tenocytes through NK-1 receptor stimulation. Br J Sports Med. 2014 Oct;48(19):1414-20.
https://doi.org/10.1136/bjsports-2013-092438 |
| 298 | Potyondy T, Uquillas JA, Tebon PJ, Byambaa B, Hasan A, Tavafoghi M, Mary H, Aninwene GE 2nd, Pountos I, Khademhosseini A, Ashammakhi N. Recent advances in 3D bioprinting of musculoskeletal tissues. Biofabrication. 2021 Mar 10;13(2):10.1088/1758-5090/abc8de.
https://doi.org/10.1088/1758-5090/abc8de |
| 299 | Peniche Silva CJ, Balmayor ER, van Griensven M. Reprogramming tendon healing: a guide to novel molecular tools. Front Bioeng Biotechnol. 2024 May 9;12:1379773.
https://doi.org/10.3389/fbioe.2024.1379773 |
| 300 | Liang W, Zhou C, Deng Y, Fu L, Zhao J, Long H, Ming W, Shang J, Zeng B. The current status of various preclinical therapeutic approaches for tendon repair. Ann Med. 2024 Dec;56(1):2337871.
https://doi.org/10.1080/07853890.2024.2337871 |
| 301 | Stephens AD, Banigan EJ, Marko JF. Chromatin's physical properties shape the nucleus and its functions. Curr Opin Cell Biol. 2019 Jun;58:76-84.
https://doi.org/10.1016/j.ceb.2019.02.006 |
| 302 | Hinz B, Lagares D. Evasion of apoptosis by myofibroblasts: a hallmark of fibrotic diseases. Nat Rev Rheumatol. 2020 Jan;16(1):11-31.
https://doi.org/10.1038/s41584-019-0324-5 |
| 303 | Morrish RB, Hermes M, Metz J, Stone N, Pagliara S, Chahwan R, Palombo F. Single Cell Imaging of Nuclear Architecture Changes. Front Cell Dev Biol. 2019 Jul 24;7:141.
https://doi.org/10.3389/fcell.2019.00141 |
| 304 | Lipp SN, Jacobson KR, Colling HA, Tuttle TG, Miles DT, McCreery KP, Calve S. Mechanical loading is required for initiation of extracellular matrix deposition at the developing murine myotendinous junction. Matrix Biol. 2023 Feb;116:28-48.
https://doi.org/10.1016/j.matbio.2023.01.003 |
| 305 | Anderson DM, George R, Noyes MB, Rowton M, Liu W, Jiang R, Wolfe SA, Wilson-Rawls J, Rawls A. Characterization of the DNA-binding properties of the Mohawk homeobox transcription factor. J Biol Chem. 2012 Oct 12;287(42):35351-35359.
https://doi.org/10.1074/jbc.M112.399386 |
| 306 | Subramanian A, Kanzaki LF, Galloway JL, Schilling TF. Mechanical force regulates tendon extracellular matrix organization and tenocyte morphogenesis through TGFbeta signaling. Elife. 2018 Nov 26;7:e38069.
https://doi.org/10.7554/eLife.38069 |
| 307 | Liu H, Zhang M, Shi M, Zhang T, Lu W, Yang S, Cui Q, Li Z. Adipose-derived mesenchymal stromal cell-derived exosomes promote tendon healing by activating both SMAD1/5/9 and SMAD2/3. Stem Cell Res Ther. 2021 Jun 10;12(1):338.
https://doi.org/10.1186/s13287-021-02410-w |
| 308 | Milz S, Benjamin M, Putz R. Molecular parameters indicating adaptation to mechanical stress in fibrous connective tissue. Adv Anat Embryol Cell Biol. 2005;178:1-71.
|
| 309 | Burd NA, West DW, Churchward-Venne TA, Mitchell CJ. Growing collagen, not muscle, with weightlifting and 'growth' hormone. J Physiol. 2010 Feb 1;588(Pt 3):395-6.
https://doi.org/10.1113/jphysiol.2009.185306 |
| 310 | Weinbaum JS, Schmidt JB, Tranquillo RT. Combating Adaptation to Cyclic Stretching By Prolonging Activation of Extracellular Signal-Regulated Kinase. Cell Mol Bioeng. 2013 Sep;6(3):279-286.
https://doi.org/10.1007/s12195-013-0289-4 |
| 311 | Erisken C, Zhang X, Moffat KL, Levine WN, Lu HH. Scaffold fiber diameter regulates human tendon fibroblast growth and differentiation. Tissue Eng Part A. 2013 Feb;19(3-4):519-28.
https://doi.org/10.1089/ten.tea.2012.0072 |
| 312 | Kew SJ, Gwynne JH, Enea D, Abu-Rub M, Pandit A, Zeugolis D, Brooks RA, Rushton N, Best SM, Cameron RE. Regeneration and repair of tendon and ligament tissue using collagen fibre biomaterials. Acta Biomater. 2011 Sep;7(9):3237-47.
https://doi.org/10.1016/j.actbio.2011.06.002 |
| 313 | Maffulli N, Sharma P, Luscombe KL. Achilles tendinopathy: Aetiology and management. J R Soc Med 2004;97(10):472-476.
https://doi.org/10.1177/0141076809701004 |
| 314 | Theodossiou SK, Murray JB, Schiele NR. Cell-cell junctions in developing and adult tendons. Tissue Barriers. 2020;8(1):1695491.
https://doi.org/10.1080/21688370.2019.1695491 |
| 315 | Lee G, Han SB, Kim SH, Jeong S, Kim DH. Stretching of porous poly (l-lactide-co-ε-caprolactone) membranes regulates the differentiation of mesenchymal stem cells. Front Cell Dev Biol. 2024 Jan 25;12:1303688.
https://doi.org/10.3389/fcell.2024.1303688 |
| 316 | Freedman BR, Bade ND, Riggin CN, Zhang S, Haines PG, Ong KL, Janmey PA. The (dys)functional extracellular matrix. Biochim Biophys Acta. 2015 Nov;1853(11 Pt B):3153-64.
https://doi.org/10.1016/j.bbamcr.2015.04.015 |
| 317 | Killian ML. Growth and mechanobiology of the tendon-bone enthesis. Semin Cell Dev Biol. 2022 Mar;123:64-73.
https://doi.org/10.1016/j.semcdb.2021.07.015 |
| 318 | Caliari SR, Weisgerber DW, Grier WK, Mahmassani Z, Boppart MD, Harley BA. Collagen Scaffolds Incorporating Coincident Gradations of Instructive Structural and Biochemical Cues for Osteotendinous Junction Engineering. Adv Healthc Mater. 2015 Apr 22;4(6):831-7.
https://doi.org/10.1002/adhm.201400809 |
| 319 | Makuku R, Werthel JD, Zanjani LO, Nabian MH, Tantuoyir MM. New frontiers of tendon augmentation technology in tissue engineering and regenerative medicine: a concise literature review. J Int Med Res. 2022 Aug;50(8):3000605221117212.
https://doi.org/10.1177/03000605221117212 |
| 320 | Costa GG, Perelli S, Grassi A, Russo A, Zaffagnini S, Monllau JC. Minimizing the risk of graft failure after anterior cruciate ligament reconstruction in athletes. A narrative review of the current evidence. J Exp Orthop. 2022 Mar 15;9(1):26.
https://doi.org/10.1186/s40634-022-00461-3 |
| 321 | Kovacevic D, Fox AJ, Bedi A, Ying L, Deng XH, Warren RF, Rodeo SA. Calcium-phosphate matrix with or without TGF-β3 improves tendon-bone healing after rotator cuff repair. Am J Sports Med. 2011 Apr;39(4):811-9.
https://doi.org/10.1177/0363546511399378 |
| 322 | Altman GH, Horan RL, Martin I, et al. Cell differentiation by mechanical stress. FASEB J 2002;16(2):270-272.
https://doi.org/10.1096/fj.01-0656fje |
| 323 | Wohlgemuth RP, Brashear SE, Smith LR. Alignment, cross linking, and beyond: a collagen architect's guide to the skeletal muscle extracellular matrix. Am J Physiol Cell Physiol. 2023 Oct 1;325(4):C1017-C1030.
https://doi.org/10.1152/ajpcell.00287.2023 |
| 324 | Grover CN, Cameron RE, Best SM. Investigating the morphological, mechanical and degradation properties of scaffolds comprising collagen, gelatin and elastin for use in soft tissue engineering. J Mech Behav Biomed Mater 2012;10:62-74.
https://doi.org/10.1016/j.jmbbm.2012.02.028 |
| 325 | Hersel U, Dahmen C, Kessler H. RGD modified polymers: Biomaterials for stimulated cell adhesion and beyond. Biomaterials 2003;24(24):4385-4415.
https://doi.org/10.1016/S0142-9612(03)00343-0 |
| 326 | Humphries JD, Byron A, Humphries MJ. Integrin ligands at a glance. J Cell Sci 2006;119(Pt 19):3901-3903.
https://doi.org/10.1242/jcs.03098 |
| 327 | Petrie TA, Raynor JE, Dumbauld DW, et al. Multivalent integrin-specific ligands enhance tissue healing and biomaterial integration. Sci Transl Med 2010;2(45):45ra60.
https://doi.org/10.1126/scitranslmed.3001002 |
| 328 | Hogan BL. Morphogenesis. Cell 1999;96(2):225-233.
https://doi.org/10.1016/S0092-8674(00)80562-0 |
| 329 | Lee K, Silva EA, Mooney DJ. Growth factor delivery-based tissue engineering: General approaches and a review of recent developments. J R Soc Interface 2011;8(55):153-170.
https://doi.org/10.1098/rsif.2010.0223 |
| 330 | Seeherman H, Wozney JM. Delivery of bone morphogenetic proteins for orthopedic tissue regeneration. Cytokine Growth Factor Rev 2005;16(3):329-345.
https://doi.org/10.1016/j.cytogfr.2005.05.001 |
| 331 | Kaiting Chen, Zezheng Liu, Xinying Zhou, Wanyu Zheng, He Cao, Zijian Yang, Zhengao Wang, Chengyun Ning, Qingchu Li, Huiyu Zhao. Hierarchy Reproduction: Multiphasic Strategies for Tendon/Ligament-Bone Junction Repair. Biomater Res. 2025;29:0132.
https://doi.org/10.34133/bmr.0132 |
| 332 | Abe S, Yamamoto M. Factors Involved in Morphogenesis in the Muscle-Tendon-Bone Complex. Int J Mol Sci. 2021 Jun 14;22(12):6365.
https://doi.org/10.3390/ijms22126365 |
| 333 | He P, Ruan D, Huang Z, Wang C, Xu Y, Cai H, Liu H, Fei Y, Heng BC, Chen W, Shen W. Comparison of Tendon Development Versus Tendon Healing and Regeneration. Front Cell Dev Biol. 2022 Jan 24;10:821667.
https://doi.org/10.3389/fcell.2022.821667 |
| 334 | Arvind V, Huang AH. Reparative and Maladaptive Inflammation in Tendon Healing. Front Bioeng Biotechnol. 2021 Jul 19;9:719047.
https://doi.org/10.3389/fbioe.2021.719047 |
| 335 | Li ZJ, Yang QQ, Zhou YL. Biological and Mechanical Factors and Epigenetic Regulation Involved in Tendon Healing. Stem Cells Int. 2023 Jan 9;2023:4387630.
https://doi.org/10.1155/2023/4387630 |
| 336 | Jiang F, Zhao H, Zhang P, Bi Y, Zhang H, Sun S, Yao Y, Zhu X, Yang F, Liu Y, Xu S, Yu T, Xiao X. Challenges in tendon-bone healing: emphasizing inflammatory modulation mechanisms and treatment. Front Endocrinol (Lausanne). 2024 Nov 6;15:1485876.
https://doi.org/10.3389/fendo.2024.1485876 |
| 337 | Tsimbouri PM. Adult Stem Cell Responses to Nanostimuli. J Funct Biomater. 2015 Jul 16;6(3):598-622.
https://doi.org/10.3390/jfb6030598 |
| 338 | Kural MH, Billiar KL. Regulating tension in three-dimensional culture environments. Exp Cell Res. 2013 Oct 1;319(16):2447-59.
https://doi.org/10.1016/j.yexcr.2013.06.019 |
| 339 | Schwartz MA. Integrins and extracellular matrix in mechanotransduction. Cold Spring Harb Perspect Biol. 2010 Dec;2(12):a005066. doi: 10.1101/cshperspect.a005066.
https://doi.org/10.1101/cshperspect.a005066 |
| 340 | Canseco JA, Kojima K, Penvose AR, Ross JD, Obokata H, Gomoll AH, Vacanti CA. Effect on ligament marker expression by direct-contact co-culture of mesenchymal stem cells and anterior cruciate ligament cells. Tissue Eng Part A. 2012 Dec;18(23-24):2549-58.
https://doi.org/10.1089/ten.tea.2012.0030 |
| 341 | Paxton JZ, Donnelly K, Keatch RP, Baar K. Engineering the bone-ligament interface using polyethylene glycol diacrylate incorporated with hydroxyapatite. Tissue Eng Part A. 2009 Jun;15(6):1201-9.
https://doi.org/10.1089/ten.tea.2008.0105 |
| 342 | Hardy M, Feehan L, Savvides G, Wong J. How controlled motion alters the biophysical properties of musculoskeletal tissue architecture. J Hand Ther. 2023 Apr-Jun;36(2):269-279.
https://doi.org/10.1016/j.jht.2022.12.003 |
| 343 | Michael M, Parsons M. New perspectives on integrin-dependent adhesions. Curr Opin Cell Biol. 2020 Apr;63:31-37.
https://doi.org/10.1016/j.ceb.2019.12.008 |
| 344 | Stuart T, Satija R. Integrative single-cell analysis. Nat Rev Genet 2019;20(5):257-272.
https://doi.org/10.1038/s41576-019-0093-7 |
| 345 | Asp M, Giacomello S, Larsson L, et al. A spatiotemporal organ-wide gene expression and cell atlas of the developing human heart. Cell 2019;179(7):1647-1660.e19.
https://doi.org/10.1016/j.cell.2019.11.025 |
| 346 | Schlaepfer DD, Mitra SK. Multiple connections link FAK to cell motility and invasion. Curr Opin Genet Dev 2004;14(1):92-101.
https://doi.org/10.1016/j.gde.2003.12.002 |
| 347 | Wu C. Focal adhesion: A focal point in current cell biology and molecular medicine. Cell Adhes Migr 2007;1(1):13-18.
https://doi.org/10.4161/cam.1.1.4081 |
| 348 | Tang Y, Zhuo D, Yu Y, Pu W, Ma Y, Zhang Y, Huang Y, Zhang Q, Tang K, Meng C, Yang D, Bai L, He D, Jin L, Zou H, Xu H, Zhu Q, Wang J, Chen Y, Liu J. Single-cell RNA sequencing reveals the CRTAC1+ population actively contributes to the pathogenesis of spinal ligament degeneration by SPP1+ macrophage. Aging Cell. 2024 Dec;23(12):e14320.
https://doi.org/10.1111/acel.14320 |
| 349 | Betzig E, Patterson GH, Sougrat R, et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science 2006;313(5793):1642-1645.
https://doi.org/10.1126/science.1127344 |
| 350 | Paz-González R, Lourido L, Calamia V, Fernández-Puente P, Quaranta P, Picchi F, Blanco FJ, Ruiz-Romero C. An Atlas of the Knee Joint Proteins and Their Role in Osteoarthritis Defined by Literature Mining. Mol Cell Proteomics. 2023 Aug;22(8):100606.
https://doi.org/10.1016/j.mcpro.2023.100606 |
| 351 | Lange JR, Fabry B. Cell and tissue mechanics in cell migration. Exp Cell Res 2013;319(16):2418-2423.
https://doi.org/10.1016/j.yexcr.2013.04.023 |
| 352 | Campagnola PJ. Second harmonic generation imaging microscopy: Applications to diseases diagnostics. Anal Chem 2011;83(9):3224-3231.
https://doi.org/10.1021/ac1032325 |
| 353 | Wang Y, Gu C, Zhao H, Li Z, Thirupathi A. Redox signaling mediated muscle atrophy in ACL injury: Role of physical exercise (Review). Mol Med Rep. 2025 May;31(5):119.
https://doi.org/10.3892/mmr.2025.13484 |
| 354 | Turčanová M, Hrtoň M, Dvořák P, Novák K, Hermanová M, Bednařík Z, Polzer S, Burša J. Full-Range Optical Imaging of Planar Collagen Fiber Orientation Using Polarized Light Microscopy. Biomed Res Int. 2021 Nov 28;2021:6879765.
https://doi.org/10.1155/2021/6879765 |
| 355 | Du J, Wu Q, Bae EJ. Epigenetics of Skeletal Muscle Atrophy. Int J Mol Sci. 2024 Jul 31;25(15):8362.
https://doi.org/10.3390/ijms25158362 |
| 356 | Avey AM, Devos F, Roberts AG, Essawy ESE, Baar K. Inhibiting JAK1, not NF-κB, reverses the effect of pro-inflammatory cytokines on engineered human ligament function. Matrix Biol. 2024 Jan;125:100-112.
https://doi.org/10.1016/j.matbio.2023.12.007 |
| 357 | Camacho DM, Collins KM, Powers RK, Costello JC, Collins JJ. Next-generation machine learning for biological networks. Cell 2018;173(7):1581-1592.
https://doi.org/10.1016/j.cell.2018.05.015 |
| 358 | Wu B, Qiu Z, Li S, Liang D, Xu J, Zhong M, Lu W, Wang D, Liu H, Zhu W, Ouyang K, Li H. [Histological properties of autogenous hamstring grafts after anterior cruciate ligament reconstruction]. Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi. 2018 Jul 15;32(7):873-879. Chinese.
|
| 359 | Onizuka N, Ito Y, Inagawa M, Nakahara H, Takada S, Lotz M, Toyama Y, Asahara H. The Mohawk homeobox transcription factor regulates the differentiation of tendons and volar plates. J Orthop Sci. 2014 Jan;19(1):172-80.
https://doi.org/10.1007/s00776-013-0485-z |
| 360 | Popov C, Burggraf M, Kreja L, Ignatius A, Schieker M, Docheva D. Mechanical stimulation of human tendon stem/progenitor cells results in upregulation of matrix proteins, integrins and MMPs, and activation of p38 and ERK1/2 kinases. BMC Mol Biol. 2015 Mar 13;16:6.
https://doi.org/10.1186/s12867-015-0036-6 |
| 361 | Ciardulli MC, Lovecchio J, Parolini O, Giordano E, Maffulli N, Della Porta G. Fibrin Scaffolds Perfused with Transforming Growth Factor-β1 as an In Vitro Model to Study Healthy and Tendinopathic Human Tendon Stem/Progenitor Cells. Int J Mol Sci. 2024 Sep 3;25(17):9563.
https://doi.org/10.3390/ijms25179563 |
| 362 | Walden G, Liao X, Donell S, Raxworthy MJ, Riley GP, Saeed A. A Clinical, Biological, and Biomaterials Perspective into Tendon Injuries and Regeneration. Tissue Eng Part B Rev. 2017 Feb;23(1):44-58.
https://doi.org/10.1089/ten.teb.2016.0181 |
| 363 | Charles MD, Christian DR, Cole BJ. The Role of Biologic Therapy in Rotator Cuff Tears and Repairs. Curr Rev Musculoskelet Med. 2018 Mar;11(1):150-161.
https://doi.org/10.1007/s12178-018-9469-0 |
| 364 | Korcari A, Nichols AEC, Buckley MR, Loiselle AE. Scleraxis-lineage cells are required for tendon homeostasis and their depletion induces an accelerated extracellular matrix aging phenotype. Elife. 2023 Jan 19;12:e84194.
https://doi.org/10.7554/eLife.84194 |
| 365 | Saraswathibhatla A, Indana D, Chaudhuri O. Cell-extracellular matrix mechanotransduction in 3D. Nat Rev Mol Cell Biol. 2023 Jul;24(7):495-516.
https://doi.org/10.1038/s41580-023-00583-1 |
| 366 | Guilluy C, Burridge K. Nuclear mechanotransduction: forcing the nucleus to respond. Nucleus. 2015;6(1):19-22.
https://doi.org/10.1080/19491034.2014.1001705 |
| 367 | Hart DA. Regulation of Joint Tissues and Joint Function: Is There Potential for Lessons to Be Learned Regarding Regulatory Control from Joint Hypermobility Syndromes? International Journal of Molecular Sciences. 2025; 26(3):1256.
https://doi.org/10.3390/ijms26031256 |
| 368 | Finni T, Vanwanseele B. Towards modern understanding of the Achilles tendon properties in human movement research. J Biomech. 2023 May;152:111583.
https://doi.org/10.1016/j.jbiomech.2023.111583 |
| 369 | Silver FH, Kelkar N, Deshmukh T. Molecular Basis for Mechanical Properties of ECMs: Proposed Role of Fibrillar Collagen and Proteoglycans in Tissue Biomechanics. Biomolecules. 2021; 11(7):1018.
https://doi.org/10.3390/biom11071018 |
| 370 | Maganaris CN, Paul JP. In vivo human tendon mechanical properties. J Physiol. 1999 Nov 15;521 Pt 1(Pt 1):307-13.
https://doi.org/10.1111/j.1469-7793.1999.00307.x |
| 371 | Graça AL, Domingues RMA, Calejo I, Gómez-Florit M, Gomes ME. Therapeutic Effects of Platelet-Derived Extracellular Vesicles in a Bioengineered Tendon Disease Model. International Journal of Molecular Sciences. 2022; 23(6):2948.
https://doi.org/10.3390/ijms23062948 |
| 372 | Shadwick RE. (1990). Elastic energy storage in tendons: mechanical differences related to function and age. Journal of Applied Physiology, 68(3): 1033-1040.
https://doi.org/10.1152/jappl.1990.68.3.1033 |
| 373 | Balestri W, Morris RH, Hunt JA, Reinwald Y. Current Advances on the Regeneration of Musculoskeletal Interfaces. Tissue Eng Part B Rev. 2021 Dec;27(6):548-571.
https://doi.org/10.1089/ten.teb.2020.0112 |
| 374 | Squier, K. (2025). Stretching boundaries : local and systemic influences on tendon function (T). University of British Columbia. Retrieved from https://open.library.ubc.ca/collections/ubctheses/24/items/1.0448056
|
| 375 | Donmazov S, Saruhan EN, Pekkan K, Piskin S. Review of Machine Learning Techniques in Soft Tissue Biomechanics and Biomaterials. Cardiovasc Eng Technol. 2024 Oct;15(5):522-549.
https://doi.org/10.1007/s13239-024-00737-y |
| 376 | Ristaniemi A, Regmi D, Mondal D, Torniainen J, Tanska P, Stenroth L, Finnilä MAJ, Töyräs J, Korhonen RK. Structure, composition and fibril-reinforced poroviscoelastic properties of bovine knee ligaments and patellar tendon. J R Soc Interface. 2021 Jan;18(174):20200737.
https://doi.org/10.1098/rsif.2020.0737 |
| 377 | Xiao S, Shao Y, Li B, Feng XQ. A micromechanical model of tendon and ligament with crimped fibers. J Mech Behav Biomed Mater. 2020 Dec;112:104086.
https://doi.org/10.1016/j.jmbbm.2020.104086 |
| 378 | Laurent, Cédric. (2018). Micromechanics of Ligaments and Tendons. 10.1016/B978-1-78548-208-3.50013-7.
https://doi.org/10.1016/B978-1-78548-208-3.50013-7 |
| 379 | Bianchi E, Ruggeri M, Rossi S, Vigani B, Miele D, Bonferoni MC, Sandri G, Ferrari F. Innovative Strategies in Tendon Tissue Engineering. Pharmaceutics. 2021; 13(1):89.
https://doi.org/10.3390/pharmaceutics13010089 |
| 380 | Holt NC, Mayfield DL. Muscle-tendon unit design and tuning for power enhancement, power attenuation, and reduction of metabolic cost. J Biomech. 2023 May;153:111585.
https://doi.org/10.1016/j.jbiomech.2023.111585 |
| 381 | Ye Y, Shetye SS, Birk DE, Soslowsky LJ. Regulatory Role of Collagen XI in the Establishment of Mechanical Properties of Tendons and Ligaments in Mice Is Tissue Dependent. J Biomech Eng. 2025 Jan 1;147(1):011003.
https://doi.org/10.1115/1.4066570 |
| 382 | Peffers MJ, Thorpe CT, Collins JA, Eong R, Wei TK, Screen HR, Clegg PD. Proteomic analysis reveals age-related changes in tendon matrix composition, with age- and injury-specific matrix fragmentation. J Biol Chem. 2014 Sep 12;289(37):25867-78.
https://doi.org/10.1074/jbc.M114.566554 |
| 383 | Freedman BR, Gordon JA, Soslowsky LJ. The Achilles tendon: fundamental properties and mechanisms for healing. J Orthop Res. 2014;32(1):122-131.
https://doi.org/10.11138/mltj/2014.4.2.245 |
| 384 | Matta C, Takács R, Ducza L, Ebeid RA, Choi H, Mobasheri A. Ion channels involved in inflammation and pain in osteoarthritis and related musculoskeletal disorders. Am J Physiol Cell Physiol. 2023 Jul 1;325(1):C257-C271.
https://doi.org/10.1152/ajpcell.00040.2023 |
| 385 | Ellingson AJ, Pancheri NM, Schiele NR. Regulators of collagen crosslinking in developing and adult tendons. Eur Cell Mater. 2022 Apr 5;43:130-152.
https://doi.org/10.22203/eCM.v043a11 |
| 386 | Lei L, Wen Z, Cao M, Zhang H, Ling SK, Fu BS, Qin L, Xu J, Yung PS. The emerging role of Piezo1 in the musculoskeletal system and disease. Theranostics. 2024 Jun 24;14(10):3963-3983.
https://doi.org/10.7150/thno.96959 |
| 387 | Tang Y, Wang Z, Xiang L, Zhao Z, Cui W. Functional biomaterials for tendon/ligament repair and regeneration. Regen Biomater. 2022 Sep 5;9:rbac062.
https://doi.org/10.1093/rb/rbac062 |
| 388 | Readioff R, Geraghty B, Kharaz YA, Elsheikh A, Comerford E. Proteoglycans play a role in the viscoelastic behaviour of the canine cranial cruciate ligament. Front Bioeng Biotechnol. 2022 Nov 15;10:984224.
https://doi.org/10.3389/fbioe.2022.984224 |
| 389 | Ji C, Wang Y, Wang Q, Wang A, Ali A, McCulloch CA. TRPV4 regulates β1 integrin-mediated cell-matrix adhesions and collagen remodeling. FASEB J. 2023 Jun;37(6):e22946.
https://doi.org/10.1096/fj.202300222R |
| 390 | Lujan TJ, Underwood CJ, Jacobs NT, Weiss JA. Contribution of glycosaminoglycans to viscoelastic tensile behavior of human ligament. J Appl Physiol (1985). 2009 Feb;106(2):423-31.
https://doi.org/10.1152/japplphysiol.90748.2008 |
| 391 | Franchi M, Ottani V, Stagni R, Ruggeri A. Tendon and ligament fibrillar crimps give rise to left-handed helices of collagen fibrils in both planar and helical crimps. J Anat. 2010 Mar;216(3):301-9.
https://doi.org/10.1111/j.1469-7580.2009.01188.x |
| 392 | Marturano JE, Xylas JF, Sridharan GV, Georgakoudi I, Kuo CK. Lysyl oxidase-mediated collagen crosslinks may be assessed as markers of functional properties of tendon tissue formation. Acta Biomater. 2014 Mar;10(3):1370-9.
https://doi.org/10.1016/j.actbio.2013.11.024 |
| 393 | Rumian AP, Wallace AL, Birch HL. Tendons and ligaments are anatomically distinct but overlap in molecular and morphological features--a comparative study in an ovine model. J Orthop Res. 2007 Apr;25(4):458-64.
https://doi.org/10.1002/jor.20218 |
| 394 | Suzuki A, Yabu A, Nakamura H. Advanced glycation end products in musculoskeletal system and disorders. Methods. 2022 Jul;203:179-186.
https://doi.org/10.1016/j.ymeth.2020.09.012 |
| 395 | Adamska O, Stolarczyk A, Gondek A, Maciąg B, Świderek J, Czuchaj P, Modzelewski K. Ligament Alteration in Diabetes Mellitus. J Clin Med. 2022 Sep 27;11(19):5719.
https://doi.org/10.3390/jcm11195719 |
| 396 | Mienaltowski MJ, Birk DE. Structure, physiology, and biochemistry of collagens. Adv Exp Med Biol. 2014;802:5-29.
https://doi.org/10.1007/978-94-007-7893-1_2 |
| 397 | Rashidi N, Harasymowicz NS, Savadipour A, Steward N, Tang R, Oswald S, Guilak F. PIEZO1-mediated mechanotransduction regulates collagen synthesis on nanostructured 2D and 3D models of fibrosis. Acta Biomater. 2025 Jan 24;193:242-254.
https://doi.org/10.1016/j.actbio.2024.12.034 |
| 398 | Sapudom J, Karaman S, Quartey BC, Mohamed WKE, Mahtani N, Garcia-Sabaté A, Teo J. Collagen Fibril Orientation Instructs Fibroblast Differentiation Via Cell Contractility. Adv Sci (Weinh). 2023 Aug;10(22):e2301353.
https://doi.org/10.1002/advs.202301353 |
| 399 | Fu S, Meng H, Inamdar S, Das B, Gupta H, Wang W, Thompson CL, Knight MM. Activation of TRPV4 by mechanical, osmotic or pharmaceutical stimulation is anti-inflammatory blocking IL-1β mediated articular cartilage matrix destruction. Osteoarthritis Cartilage. 2021 Jan;29(1):89-99.
https://doi.org/10.1016/j.joca.2020.08.002 |
| 400 | Thornton GM, Oliynyk A, Frank CB, Shrive NG. Ligament creep cannot be predicted from stress relaxation at low stress: a biomechanical study of the rabbit medial collateral ligament. J Orthop Res. 1997 Sep;15(5):652-6.
https://doi.org/10.1002/jor.1100150504 |
| 401 | Deymier AC, An Y, Boyle JJ, Schwartz AG, Birman V, Genin GM, Thomopoulos S, Barber AH. Micro-mechanical properties of the tendon-to-bone attachment. Acta Biomater. 2017 Jul 1;56:25-35.
https://doi.org/10.1016/j.actbio.2017.01.037 |
| 402 | Fang F, Casserly M, Robbins J, Thomopoulos S. Hedgehog signaling directs cell differentiation and plays a critical role in tendon enthesis healing. NPJ Regen Med. 2025 Jan 20;10(1):3.
https://doi.org/10.1038/s41536-025-00392-4 |
| 403 | Chen C, Shi Q, Li M, Chen Y, Zhang T, Xu Y, Liao Y, Ding S, Wang Z, Li X, Zhao C, Sun L, Hu J, Lu H. Engineering an enthesis-like graft for rotator cuff repair: An approach to fabricate highly biomimetic scaffold capable of zone-specifically releasing stem cell differentiation inducers. Bioact Mater. 2022 Jan 5;16:451-471.
https://doi.org/10.1016/j.bioactmat.2021.12.021 |
| 404 | Friese N, Gierschner MB, Schadzek P, Roger Y, Hoffmann A. Regeneration of Damaged Tendon-Bone Junctions (Entheses)-TAK1 as a Potential Node Factor. Int J Mol Sci. 2020 Jul 22;21(15):5177.
https://doi.org/10.3390/ijms21155177 |
| 405 | Knott L, Bailey AJ. Collagen cross-links in mineralizing tissues: a review of their chemistry, function, and clinical relevance. Bone. 1998 Mar;22(3):181-7.
https://doi.org/10.1016/S8756-3282(97)00279-2 |
| 406 | Roffino S, Camy C, Foucault-Bertaud A, Lamy E, Pithioux M, Chopard A. Negative impact of disuse and unloading on tendon enthesis structure and function. Life Sci Space Res (Amst). 2021 May;29:46-52
https://doi.org/10.1016/j.lssr.2021.03.001 |
| 407 | Koria L, Farndon M, Jones E, Mengoni M, Brockett C. Changes in subchondral bone morphology with osteoarthritis in the ankle. PLoS One. 2024 Jun 18;19(6):e0290914.
https://doi.org/10.1371/journal.pone.0290914 |
| 408 | Song L, Golman M, Abraham AC, Zelzer E, Thomopoulos S. A role for TGFβ signaling in Gli1+ tendon and enthesis cells. FASEB J. 2024 Mar 31;38(6):e23568.
https://doi.org/10.1096/fj.202301452R |
| 409 | Ozone K, Minegishi Y, Takahata K, Takahashi H, Yoneno M, Hattori S, Xianglan L, Oka Y, Murata K, Kanemura N. Eccentric contraction-dominant exercise leads to molecular biological changes in enthesis and enthesopathy-like morphological changes. J Orthop Res. 2023 Mar;41(3):511-523.
https://doi.org/10.1002/jor.25399 |
| 410 | Wang X, Xie L, Crane J, Zhen G, Li F, Yang P, Gao M, Deng R, Wang Y, Jia X, Fan C, Wan M, Cao X. Aberrant TGF-β activation in bone tendon insertion induces enthesopathy-like disease. J Clin Invest. 2018 Feb 1;128(2):846-860.
https://doi.org/10.1172/JCI96186 |