Combined Effects of Toxic Metals and Oxidative Stress on the Development and Health of Sea Trout (Salmo Trutta L.)
bNicolaus Copernicus University in Toruń, Collegium Medicum in Bydgoszcz, Department of Medical Biology and Biochemistry, Department of Ecology and Environmental Protection, M. Skłodowska-Curie St. 9, 85-094 Bydgoszcz, Poland,
cUniversity of Zielona Góra, Faculty of Biological Sciences, Institute of Biological Sciences, Department of Biotechnology, Prof. Z. Szafran St. 1, 65-516 Zielona Góra, Poland
Keywords
Abstract
Background/Aims:
Contaminants in the environment pose a considerable threat to biodiversity, ecological balance, and the health of both wildlife and humans, particularly through the transfer of these harmful substances via fish in the food chain.Methods:
This study focused on the developmental stages of sea trout (Salmo trutta L.) in both riverine and Baltic Sea environments, with the aim of exploring how chemical element accumulation influences oxidative stress biomarkers in these species.Results:
The findings revealed notable age- and tissue-specific patterns in the accumulation of chemical elements in sea trout. Specifically, higher levels of lead (Pb), arsenic (As), mercury (Hg), and tin (Sn) were detected in the muscle tissues of adult trout, while cadmium (Cd) primarily accumulated in the gills, particularly in smolts. These results underscore the influence of both age and tissue type on the bioaccumulation of contaminants in the trout, highlighting how the accumulation of toxic elements contributes to increased oxidative stress in the fish. This oxidative stress, reflected by increased lipid peroxidation (TBARS) and carbonyl derivatives of oxidatively modified proteins, was closely related to the presence of contaminants such as Cd, Pb, As, Hg, and Sn. Gills, which are directly exposed to waterborne pollutants, exhibited significantly higher levels of oxidative damage compared to muscle tissue, consistent with the greater accumulation of metals in this organ. Despite higher total antioxidant status (TAS) in muscle tissue, both muscle and gill tissues of adult trout showed signs of considerable oxidative stress, indicating the cumulative effects of prolonged exposure to these contaminants.Conclusion:
The study highlights the detrimental consequences of chemical element contamination on the health of trout, with a particular emphasis on oxidative damage, and calls for effective environmental management to protect aquatic species from the long-term effects of exposure to contaminants. Furthermore, the correlation and regression analysis conducted revealed significant patterns, demonstrating positive correlations between the accumulation of Cd, Pb, and As in the gills of adult trout, and between Pb and oxidative stress markers in smolts. Additionally, the analysis indicated that mercury contributes significantly to oxidative damage.Introduction
Pollutants in the environment, especially those introduced by industrial, agricultural and urban activities, pose a significant threat to biodiversity, ecological balance and the health of wildlife and humans. In aquatic ecosystems such as the Baltic Sea, these pollutants - ranging from heavy metals and persistent organic pollutants (POPs) to microplastics - can accumulate and biomagnified through the food chain [1]. Fish, both consumers and prey, are important vectors for the transfer of these pollutants to higher trophic levels, including humans. The Baltic Sea is particularly vulnerable to contamination due to its semi-enclosed geography, limited water exchange and intensive human activity in the surrounding areas. Pollutants such as mercury, cadmium, lead and arsenic often accumulate in the tissues of fish species, including Baltic Sea trout, an ecologically and economically important species [2]. These contaminants threaten not only the health of the fish populations themselves - potentially impairing growth, reproduction and immune function - but also the predators, including humans, that consume these fish [3, 4].
The Baltic Sea poses a significant environmental threat to migratory species, such as sea trout (Salmo trutta L.), through a combination of factors including pollution, habitat degradation, and climate change [1]. A major concern is the elevated levels of contaminants, including toxic metals [2], persistent organic pollutants (POPs), and various other pollutants, which accumulate in the marine ecosystem [1, 3]. These contaminants, originating mainly from industrial discharges, agricultural runoff, and sewage, pose serious risks to aquatic organisms. The bioavailability of toxic metals in the Baltic Sea may exceed that in other marine environments due to its special characteristics, such as relatively low salinity and enclosed geography [4]. As a result, migratory fish species, including sea trout, are at increased risk of metal bioaccumulation, leading to potential health problems, such as oxidative stress, impaired growth, and reproductive problems [5-7]. The vulnerability of the Baltic Sea ecosystem to overfishing, habitat destruction, and climate change exacerbates the challenges faced by migratory trout. As pollution increases and the ecosystem’s ability to support healthy populations of marine life decreases, the survival and well-being of such species as sea trout are increasingly at risk. Therefore, protecting and restoring the ecological health of the Baltic Sea is essential not only for the conservation of migratory trout, but also for the conservation of the region’s biodiversity as a whole [8].
The study of trout, in particular Salmo trutta L. or the migratory sea trout, at different life stages is essential due to the ecological importance of the species and its sensitivity to environmental changes [9]. As a key indicator species within freshwater ecosystems, migratory trout play a central role in maintaining the balance of aquatic food webs. Their presence and health can serve as a reflection of the overall quality of the aquatic environment, making them valuable for environmental monitoring and conservation efforts, particularly in response to anthropogenic stressors in both freshwater and marine habitats [10]. By studying trout throughout their developmental stages, researchers can gain comprehensive insights into the effects of environmental factors, such as pollution, temperature fluctuations, and habitat degradation on their growth, survival, and reproductive success [11].
The species in question has significant economic, ecological, and cultural importance. Trout serve as an important species for both recreational and commercial fisheries, contributing to the livelihoods of communities and supporting local economies, as demonstrated in the study by Lynch et al . [12]. Furthermore, their ability to thrive in a wide range of freshwater and marine habitats positions them as an important component of biodiversity in river systems and coastal environments. As a keystone species, trout influence populations of other aquatic organisms, making monitoring their health and developmental stages essential for the conservation of aquatic ecosystems [13]. However, trout are highly sensitive to environmental changes, making them vulnerable to habitat degradation, water pollution and climate change [14]. Their migratory nature exposes them to a variety of ecosystems, each potentially affected by human activities and environmental changes [10]. Variations in water temperature, pollution levels, and river modifications can disrupt their migratory patterns, reproductive cycles, and overall health. This makes trout an exemplary species for studying the effects of environmental change and highlights the need for targeted conservation efforts to protect them from the ongoing challenges posed by climate change and anthropogenic activities [15].
The developmental stages of migrating trout (Salmo trutta L.) in riverine and Baltic Sea environments are markedly influenced by the interaction between toxic metals and oxidative stress parameters [7, 16], as depicted in Fig. 1. In freshwater conditions in rivers, trout show different physiological and biochemical responses than in the saline environment of the Baltic Sea. Freshwater ecosystems are often exposed to elevated concentrations of pollutants, including toxic metals, which can accumulate in trout tissues, especially during critical developmental stages, such as the juvenile and smolt phases [17, 18]. The presence of these metals in rivers induces oxidative stress as the trout’s immune system attempts to mitigate the harmful effects of these toxins, resulting in changes in such stress biomarkers as 2-thiobarbituric acid reactive substances (TBARS), a common indicator of oxidative-induced lipid damage [16, 19-21].

Fig. 1: Sea trout lifecycle (egg, parr, smolt, adult, spawner).
When trout migrate from freshwater rivers to the Baltic Sea, they encounter a wide range of environmental stressors [22]. In the marine environment, variations in salinity and the presence of additional contaminants can affect the ability of trout to regulate metal accumulation in their tissues. The transition from freshwater to saltwater can significantly affect the metabolic processes and stress response mechanisms of fish. In saltwater, the bioavailability and toxicity of certain metals may change due to variations in pH, salinity, and organic matter content, which may affect the interactions between metals and the biological systems of the fish [22]. Consequently, oxidative stress parameters may differ between these two environments as the fish body adapts to the different environmental conditions.
The main objective of these studies is to improve understanding of the effects of various environmental factors, including water quality, pollution and habitat conditions, on the health, development, and survival of migrating sea trout (Salmo trutta L.) at different life stages [17]. Understanding their responses to environmental contaminants is essential for broader environmental management practices [10, 23].
Numerous studies have highlighted the effects of chemical elements, such as cadmium (Cd), lead (Pb), mercury (Hg), and zinc (Zn), on oxidative stress processes in some tissues of sea trout, thereby disrupting cellular homeostasis and metabolism [16, 24]. These metals catalyse the production of reactive oxygen species (ROS) through Fenton-like reactions and mitochondrial interference, while inhibiting enzymatic antioxidants, such as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx) [21, 25]. They also deplete non-enzymatic antioxidants, such as glutathione (GSH), resulting in reduced defence against oxidative damage. The resulting ROS induce lipid peroxidation, protein oxidation, and DNA damage, leading to membrane disruption, structural protein changes, and genomic instability. Together, these effects impair growth, immune response, and reproduction, ultimately jeopardising the health and fitness of sea trout populations [26].
Furthermore, the interaction between toxic metals and oxidative stress parameters may be exacerbated during certain developmental stages, as previously indicated by Saç and Yeltekin [27]. For example, during the early juvenile stages in rivers, when trout are more vulnerable and have less developed detoxification mechanisms, the accumulation of such metals as Cd, Pb, and As can lead to increased levels of oxidative stress [28-31]. In contrast, adult trout in the marine environment, especially after migration, may have more advanced detoxification systems, which may mitigate the harmful effects of these metals to some extent. However, the long-term accumulation of metals during migration may still lead to increased oxidative damage, especially during periods of environmental stress [32, 33], such as elevated water temperatures or reduced oxygen levels in the Baltic Sea. A study by the authors [34] analysed the mineral composition and heavy metal concentrations in the muscles of Baltic Sea trout. The study showed that the levels of toxic elements such as arsenic, lead, cadmium and mercury were below international safety limits. The results indicate that Baltic Sea trout accumulate beneficial minerals such as copper and magnesium while maintaining low levels of harmful metals, highlighting their environmental and ecological importance.
The interaction between metals and oxidative stress parameters in trout is likely to be influenced by a combination of biological and environmental factors, with the developmental stage and the specific conditions of riverine and Baltic Sea environments playing a crucial role [16], as shown in Fig. 2. The riverine environment may result in greater accumulation of toxic metals, leading to more pronounced oxidative stress during certain life stages. Conversely, the transition to the Baltic Sea presents additional challenges, such as changes in salinity and variations in metal bioavailability, which may affect the oxidative stress response [18, 21, 23]. Understanding these interactions is critical for assessing the overall health of migratory trout populations and developing effective conservation strategies to protect them from the detrimental effects of pollution in both freshwater and marine habitats [7].
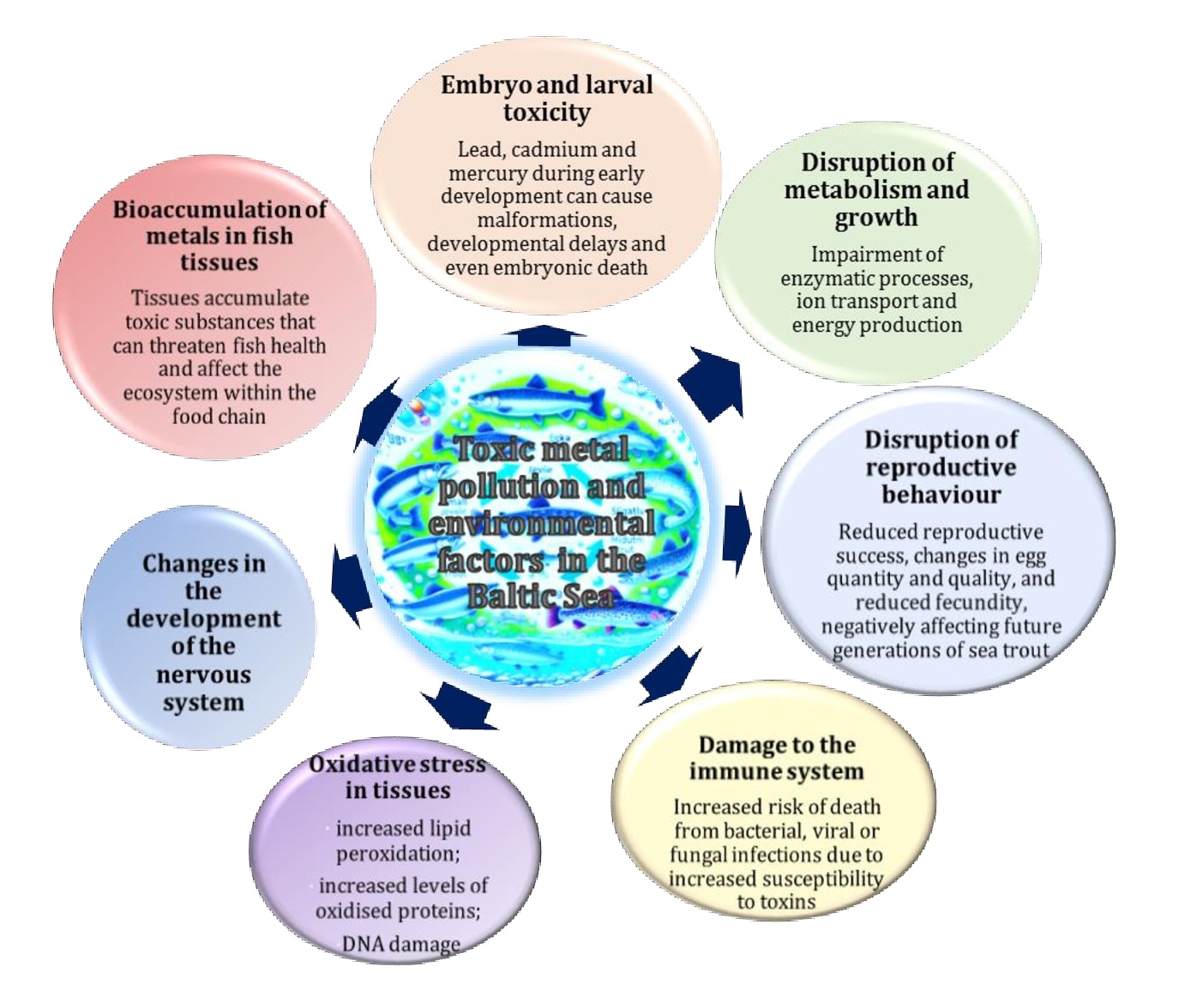
Fig. 2: Influence of toxic element pollution and environmental factors on the development of sea trout in the Baltic Sea. The unique characteristics of the Baltic Sea, including its low water exchange rate and low salinity coupled with the presence of chemical elements, pose a significant threat to the life cycle of sea trout. Metal contamination in this ecosystem affects the health and survival of fish at all developmental stages, resulting in impaired immunity, reproductive disorders and bioaccumulation of toxins in fish tissues. These effects have long-term consequences for entire sea trout populations.
A key aim of this research is to assess the adaptation of migratory trout to different aquatic environments, such as freshwater rivers and saltwater seas, and the effects of these environments on contaminant accumulation and oxidative stress [34]. By comparing these conditions, the research can elucidate the specific vulnerabilities of trout in both types of habitat, helping to refine conservation measures that encompass the full migratory life cycle of the species [22, 35]. In addition, this research may contribute to a broader understanding of the impact of anthropogenic activities, such as industrial pollution, agricultural run-off, and climate-induced changes in water temperature, on aquatic ecosystems [36]. The findings may have wider implications for biodiversity conservation and the protection of other aquatic species that share similar ecological niches [1, 5]. By establishing links between environmental conditions and the health of migratory trout, the study can provide valuable data for regulators, researchers, and conservation organisations to implement more effective measures for sustainable water management, pollution control, and ecosystem restoration. Ultimately, these studies aim to contribute to the conservation of both migratory trout and the aquatic ecosystems that support them, ensuring their long-term viability in the face of the ongoing environmental challenges.
The aim of this study was to thoroughly analyse the effects of chemical elements on oxidative stress in sea trout (Salmo trutta L.) at different developmental stages (smolts and adults) and in different tissues (muscle and gills) in order to obtain a comprehensive understanding of the effects of environmental pollution on the health of trout in terms of changes in metabolism and tissue function, which is crucial for the protection of this species. The study focused on four main aspects: (1) measurement of the levels of chemical elements such as Cd, Pb, As, Hg, and Sn in different tissues and at different developmental stages; (2) assessment of the level of oxidative stress determined by such indicators as lipid peroxidation (TBARS) and oxidative protein modifications; (3) MANOVA statistical analysis to evaluate interactions between variables and the influence of metals on oxidative stress as a function of developmental stage; and (4) detailed correlation and regression analyses to determine the relationships between metal accumulation and oxidative stress markers and to identify significant associations between these factors.
Materials and Methods
Study area and fish collection.
The Słupia River, located in the Pomeranian region of northern Poland, represents an important river system for ichthyological activities in Central Pomerania. Classified as a coastal river, its entire catchment area lies within the Pomeranian Voivodeship. The middle section of the river flows through the Polanowska Upland, while the lower section flows through the Damnica Upland, the Słupsk Plain, and the Słowiński Coast, finally flowing into the Baltic Sea in Ustka. Samples of migrating trout were systematically collected from various points along the Słupia and its tributaries, as shown in Fig. 3.
The analysis showed that the water quality of the Słupia River in the Słupsk region complies with the European standard EN ISO 8689-2 and the Water Framework Directive, achieving a classification of first class purity, as shown by Obolewski [37]. Smolt-stage trout samples were collected from several Pomeranian rivers, including the Glaźna, Skotawa, Kamienna, and Kwacza near Słupsk city (Fig. 3). Smolt sampling was conducted using electrofishing equipment powered by a generator with a DC adaptor. Adult trout were sampled at the mouth of the Słupia River in Ustka (Pomeranian Voivodeship, northern Poland). A total of 69 adult and 128 smolt trout samples were used for this analysis. This research was conducted from 2018 to 2021 in cooperation with the ‘Dolina Słupia’ Landscape Park and the Słupsk District Office of the Polish Angling Association under the environmental permit DROS.AR.MW.6052-16/10.
Our study follows the terminology and definitions previously explored by Pratten and Shearer (1983) [38], who established that sea trout are anadromous while brown trout are the resident form of the same species. We studied two developmental stages of trout: the smolt (juvenile) and the adult (Fig. 3). It is well documented that during the smolt stage, trout reach a size of 10-20 cm and are characterised by a transitional habitat stage, as the fish migrate to the sea [18]. The adult stage of sea trout includes individuals that return to rivers as mature specimens after spending 2-3 years at sea. These fish typically weigh 2-3 kg, are sexually mature, and have a silver colouration as described in studies [39, 40]. The effect of sex on metal content was not investigated in this analysis. The relationship between developmental stages and oxidative stress parameters as well as biochemical characteristics in these conditions has been presented in our previous studies [16, 21].
The fish were euthanised by percussion stunning followed by crushing of the brain. Tissue samples were collected immediately, frozen on site at –80°C, and then homogenised in the laboratory for analysis.
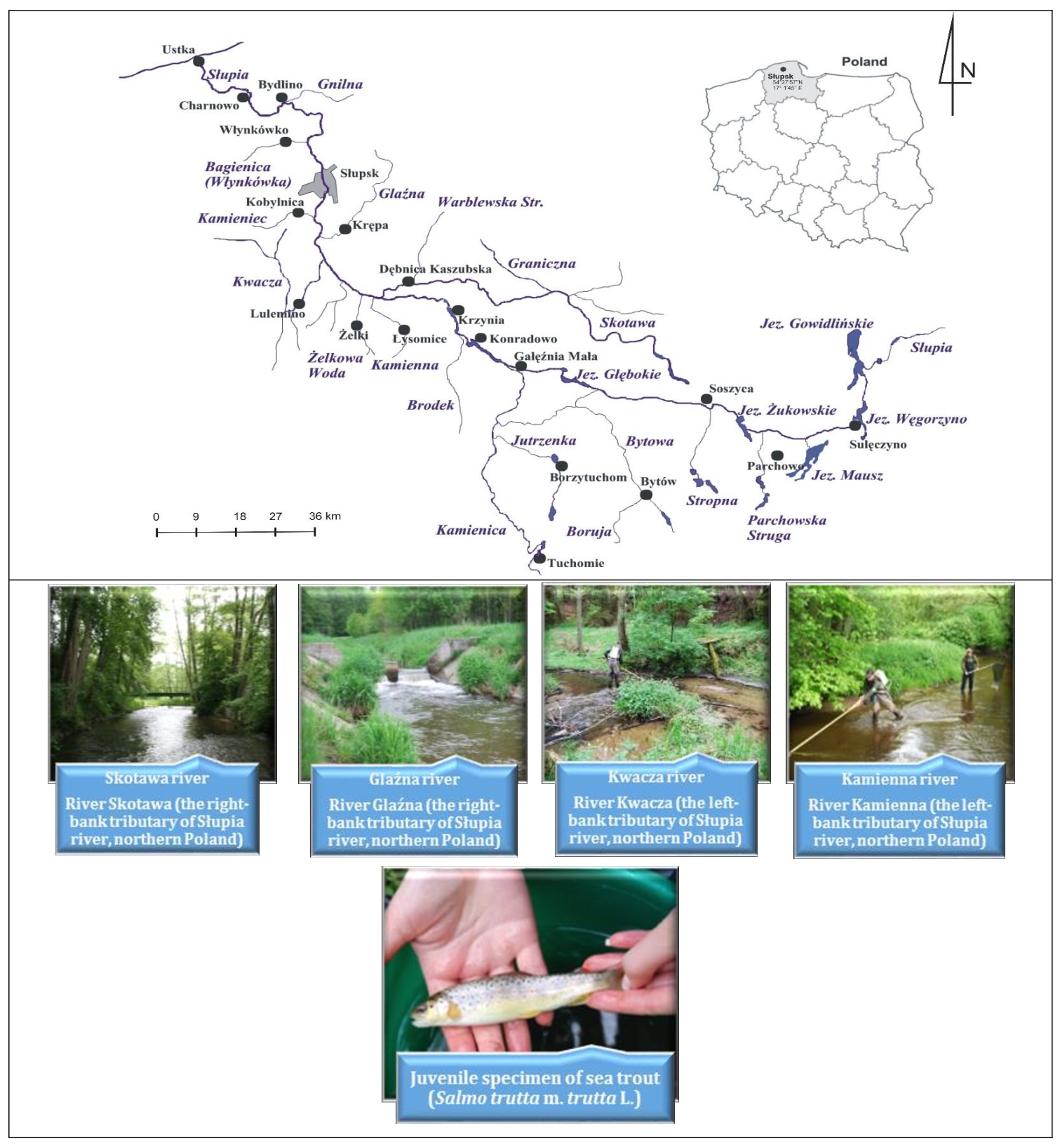
Fig. 3: Maps of the Pomeranian region of northern Poland showing the location of the town of Słupsk. This is the area where trout samples were collected for the study. The fish were caught in four tributaries of the Słupia River: Glaźna, Skotawa, Kamienna, and Kwacza (Central Pomerania, northern Poland). Photo Natalia Kurhaluk.
Tissue collection and preparation. Gill and muscle tissues were collected from both smolt and adult stages of the fish for the biochemical analysis of oxidative stress biomarkers. Muscle tissue samples were collected specifically from the dorsal region above the lateral line. The collected tissues were homogenised in chilled 0.1 M Tris-HCl buffer (pH 7.2) at a ratio of 1:9 (tissue to buffer). The resulting homogenate was centrifuged at 3, 000 rpm for 15 min at 4°C and used as the starting material for subsequent biochemical analyses.
Quantitative analysis of chemical elements
The digestion of the sample was conducted in accordance with the following procedure. Dried animal tissues were digested using concentrated nitric acid (8 ml, 69–70%) and hydrogen peroxide (2 ml, 30%) via microwave digestion. The digestion program included sequential heating: Five minutes at 190°C (a 25-minute ramp), five minutes at 200°C (a 5-minute ramp), and five minutes at 210°C (a 5-minute ramp). The resulting solutions were then diluted to 50 ml with 0.05 μS/cm deionised water.
The instruments and reagents used are listed below. Trace element analysis was performed using an Agilent 7500ce ICP-MS with a micromist nebuliser, Peltier-cooled double-pass spray chamber, and a peristaltic pump. The system featured a shielding torch, off-axis ion lenses, and an Octupole Reaction System (ORS) chamber with hydrogen and helium (6.0, 99.9999%) to reduce interference. A quadrupole mass analyser with hyperbolic rods and a dual-mode detector (digital/analogue) provided a dynamic range extending over nine orders of magnitude. High-purity argon (5.0, 99.999%) was used as the carrier gas. The utilisation of internal standards (45Sc, 89Y, 159Tb) was instrumental in minimising matrix effects and ensuring the stability of measurements. Quality control measures were implemented to ensure the reliability of the analytical process. Each batch included blanks and a certified reference material (NCS ZC73016 chicken). The measurement uncertainty was 10%, with recoveries ranging from 90% to 110%.
Analyses of biomarkers of oxidative stress
The results are expressed as mean ± S.E.M., which provides a clear measure of central tendency and variability. Where the data did not follow a normal distribution, a logarithmic transformation was performed to meet the assumptions of parametric statistical tests. Pearson’s regression analysis was used within the multiple regression module to examine relationships between parametric variables, including the calculation of correlation coefficients (r), regression equations, and the statistical significance of these relationships (P values). This approach elucidated the relationship between oxidative stress biomarkers and elemental concentrations.
The arithmetic mean concentrations of oxidative stress biomarkers and the elemental content in trout tissues (muscle and gills) were compared between developmental stages (smolt and adult) using two-way ANOVA. Multivariate significance tests and two-way ANOVA allowed multiple factors to be assessed simultaneously. The regression and correlation analyses provided insight into the relationships between chemical elements, oxidative stress biomarkers, and tissue-specific responses at different developmental stages of trout. This method facilitated the evaluation of the effects of chemical elements, such as Cd, Pb, As, Hg, and Sn, on the developmental stage and tissue type. The significance of these effects was further assessed using multivariate testing to ensure robustness and reliability in the interpretation of results.
A two-way classification model was used to assess the combined effects of chemical elements on the developmental stage and tissue type parameters. The full model was described using multiple correlation coefficients (R), coefficient of determination (R²), and adjusted R² (R²adj) to account for random error in data analysis. These metrics provided insight into the proportion of variability explained by the model. The proportion of variance explained by oxidative stress biomarkers and biochemical parameters was assessed using the sum of squares (SS) test. The F-test was used to determine the significance of these proportions, providing a comprehensive understanding of the relationships and effects investigated [46].
Results
The accumulation of chemical elements, including Cd, Pb, As, Hg, and Sn, in the muscle tissue and gills of sea trout was analysed and the results are shown in Figures 4 and 5. Our results showed distinct patterns of metal accumulation influenced by both the tissue type and the age group.
Cadmium (Cd) levels were significantly higher in gill tissue of adult sea trout compared to muscle tissue, indicating that the gills are a primary site for metal uptake (Fig. 4A). Interestingly, juvenile trout exhibited higher overall Cd accumulation than adults, which may reflect developmental differences in metabolism or exposure history. As shown in Fig. 4A, cadmium accumulation in gill tissues was significantly higher in adults than in smolts, whereas the opposite trend was observed in muscle tissues, where smolts exhibited higher Cd levels than adults. These results highlight age- and tissue-specific differences in Cd uptake and detoxification. In adult trout, elevated gill Cd levels may reflect prolonged exposure in the marine environment, where gill tissue, as the primary site of ion exchange, is particularly susceptible to metal accumulation. Such bioaccumulation could impair osmoregulatory function by disrupting ion transport mechanisms. In contrast, the elevated Cd concentrations observed in the muscle tissue of smolts may indicate a limited detoxification capacity at this developmental stage, resulting in greater systemic distribution and storage of the metal. Taken together, these results highlight the physiological susceptibility of different life stages to cadmium exposure and emphasise the importance of tissue-specific analysis in ecotoxicological assessments.
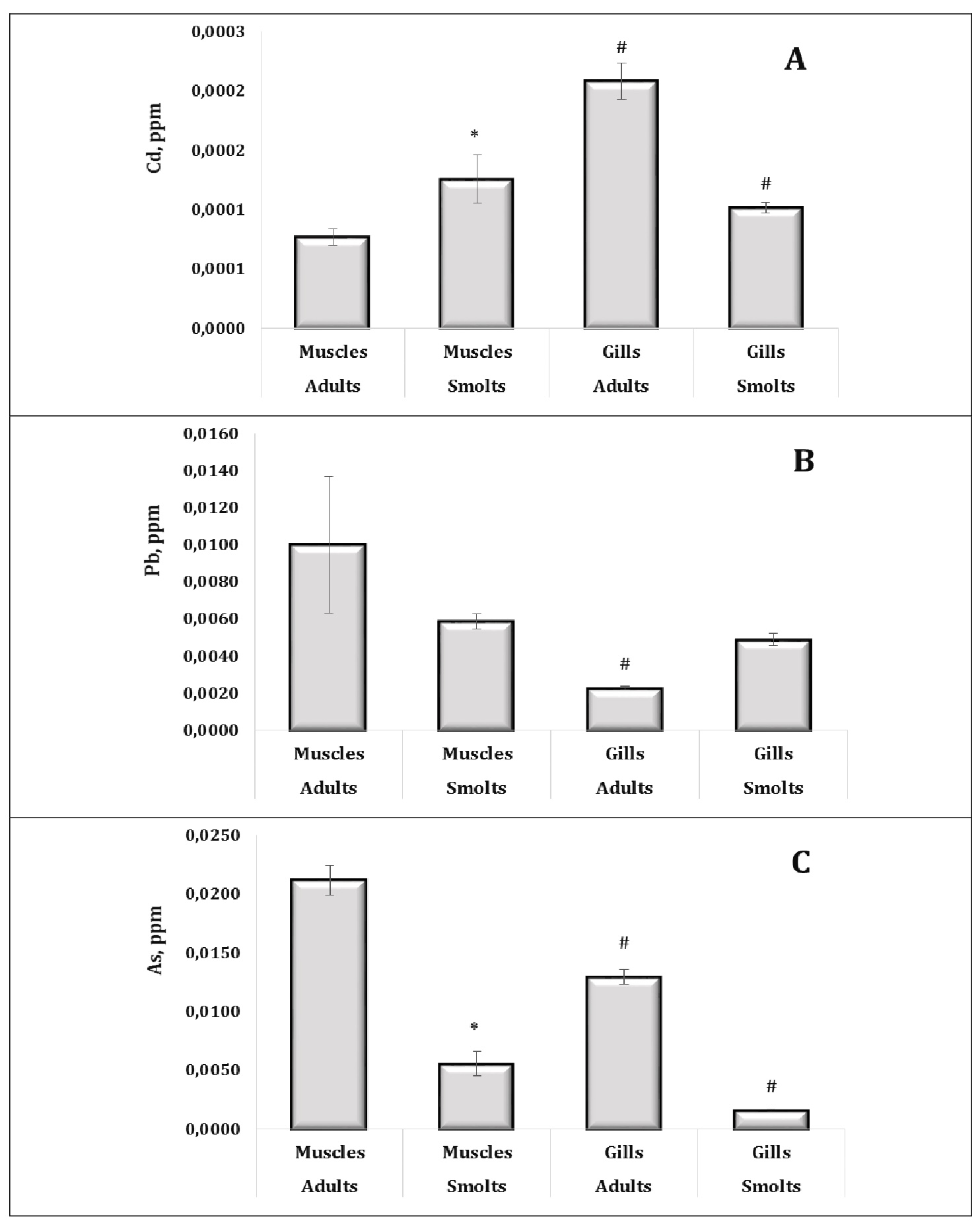
Fig. 4: Contents of Cd (A, ppm), Pb (B, ppm), and As (C, ppm) in muscle and gill tissues of adult and juvenile sea trout (Salmo trutta L.) sampled from the Pomeranian region (northern Poland). * Changes are significant (p < 0.05) for one tissue type and different age groups; # Changes are significant (p < 0.05) for one age group and different tissue types. Results are expressed as mean ± S.E.M. (adults, muscles, n = 69; smolts, muscles, n = 128; adults, gills, n = 50; smolts, gills, n = 60). Differences between groups were analysed using two-way ANOVA followed by Bonferroni's post-hoc test.
For lead and arsenic, adult muscle tissue showed a greater propensity to uptake these elements, with statistically significant differences supporting these observations (Fig. 4B, 4C). In addition, arsenic showed a similar dependence in the gills, indicating its dual affinity for both muscle and gill tissues in adult trout (Fig. 4C).
A consistent pattern was observed for mercury, where higher concentrations were recorded in muscle tissue compared to gills in both age groups, highlighting muscle tissue as a primary storage site for Hg (Fig. 5A). This trend suggests that Hg bioaccumulation continues throughout the life cycle of the fish. Furthermore, the tin levels were significantly higher in the muscle tissue of adult trout compared to smolts, suggesting a cumulative effect of exposure over time (Fig. 5B). These results highlight the differences in the uptake and storage of chemical elements between tissues and age groups, reflecting the complex interplay between environmental exposure, physiological processes, and tissue-specific retention mechanisms.
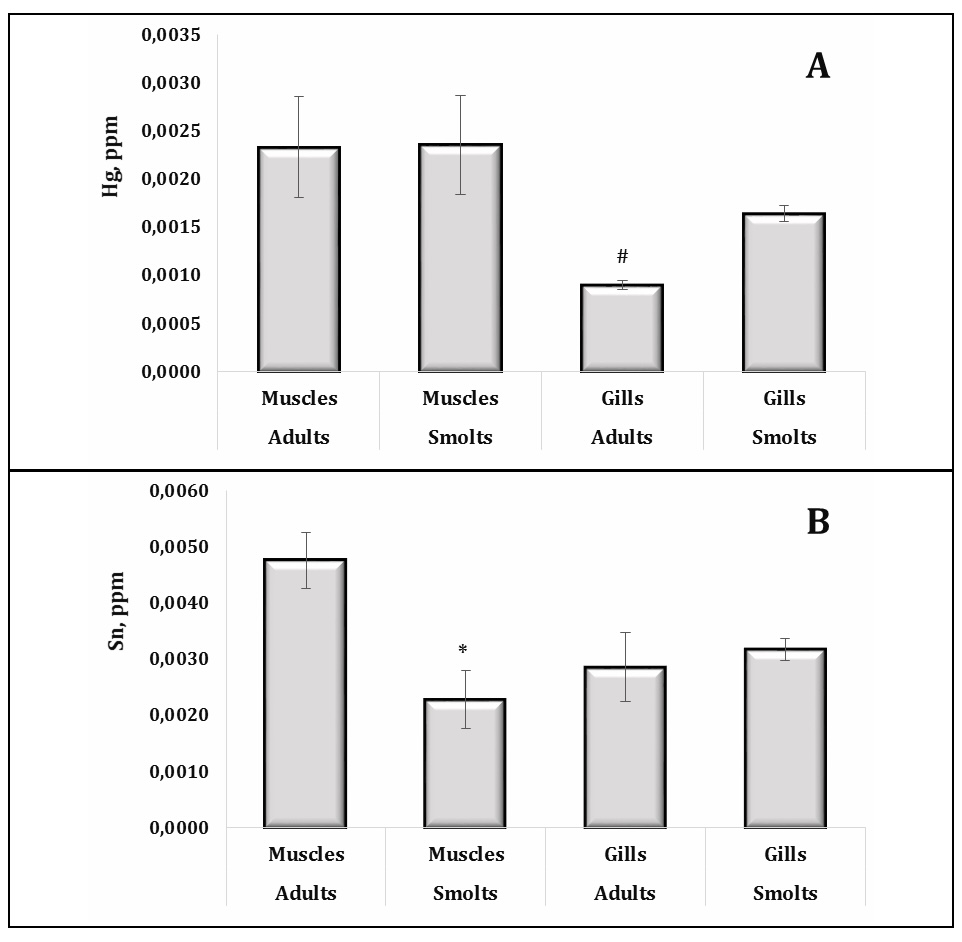
Fig. 5: Content of Hg (A, ppm) and Sn (B, ppm) in muscle and gill tissues of adult and juvenile sea trout (Salmo trutta L.) sampled from the Pomeranian region (northern Poland). * Changes are significant (p < 0.05) for one tissue type and different age groups; # Changes are significant (p < 0.05) for one age group and different tissue types. Results are expressed as mean ± S.E.M. (adults, muscles, n = 69; smolts, muscles, n = 128; adults, gills, n = 50; smolts, gills, n = 60). Differences between groups were analysed using two-way ANOVA followed by Bonferroni's post-hoc test.
Thus, our results indicate that chemical element accumulation in sea trout increases with age and prolonged exposure to contaminated environments, with muscle tissue serving as a primary storage site for persistent contaminants, such as cadmium and mercury. In addition, the gills, which are directly exposed to waterborne metals, play a critical role in the initial uptake of contaminants, such as lead and arsenic, highlighting the combined effects of chronic and acute contamination on trout health.
Our results showed that lipid peroxidation, as assessed by TBARS levels, was significantly higher in adult trout compared to smolts, reflecting increased oxidative stress with age and environmental exposure (Fig. 6A). Notably, the gills had higher TBARS levels than muscle tissue in adult individuals, suggesting that gills are more susceptible to oxidative damage due to their direct interaction with waterborne pollutants. This pattern highlights the critical role of gills as a frontline organ exposed to environmental stressors, particularly in polluted habitats.
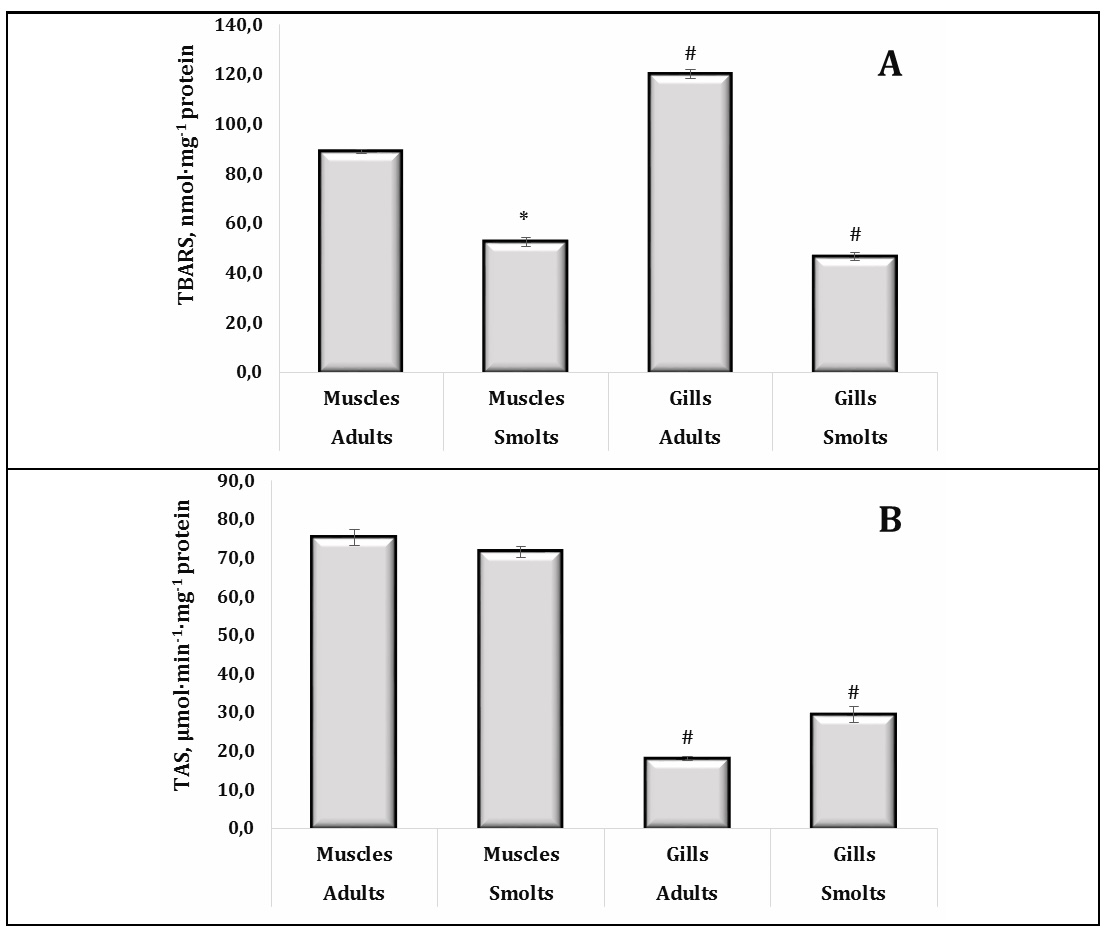
Fig. 6: TBARS levels (A, nmol·mg-1 protein) and total antioxidant status (B, μmol·min-1·mg-1 protein) in muscle and gill tissues of adult and juvenile sea trout (Salmo trutta L.) sampled from the Pomeranian region (northern Poland). * Changes are significant (p < 0.05) for one tissue type and different age groups; # Changes are significant (p < 0.05) for one age group and different tissue types. Results are expressed as mean ± S.E.M. (adults, muscles, n = 69; smolts, muscles, n = 128; adults, gills, n = 50; smolts, gills, n = 60). Differences between groups were analysed using two-way ANOVA followed by Bonferroni's post-hoc test.
However, the total antioxidant status (TAS) in muscle tissue remained consistent across the developmental stages, indicating a stable antioxidant defence mechanism in this tissue (Fig. 6B). Furthermore, the TAS levels were higher in muscle than in gills, highlighting the enhanced ability of muscle tissue to counteract oxidative damage. Oxidative protein modifications (OMP AD and OMP KD) were found to be more pronounced in adults, with muscle tissues showing higher levels of these modifications than gills (Fig. 7A, 7B). This suggests a cumulative effect of oxidative stress over time, particularly in metabolically active tissues, such as muscle, and further emphasises the impact of ageing and environmental exposure on oxidative damage in trout.
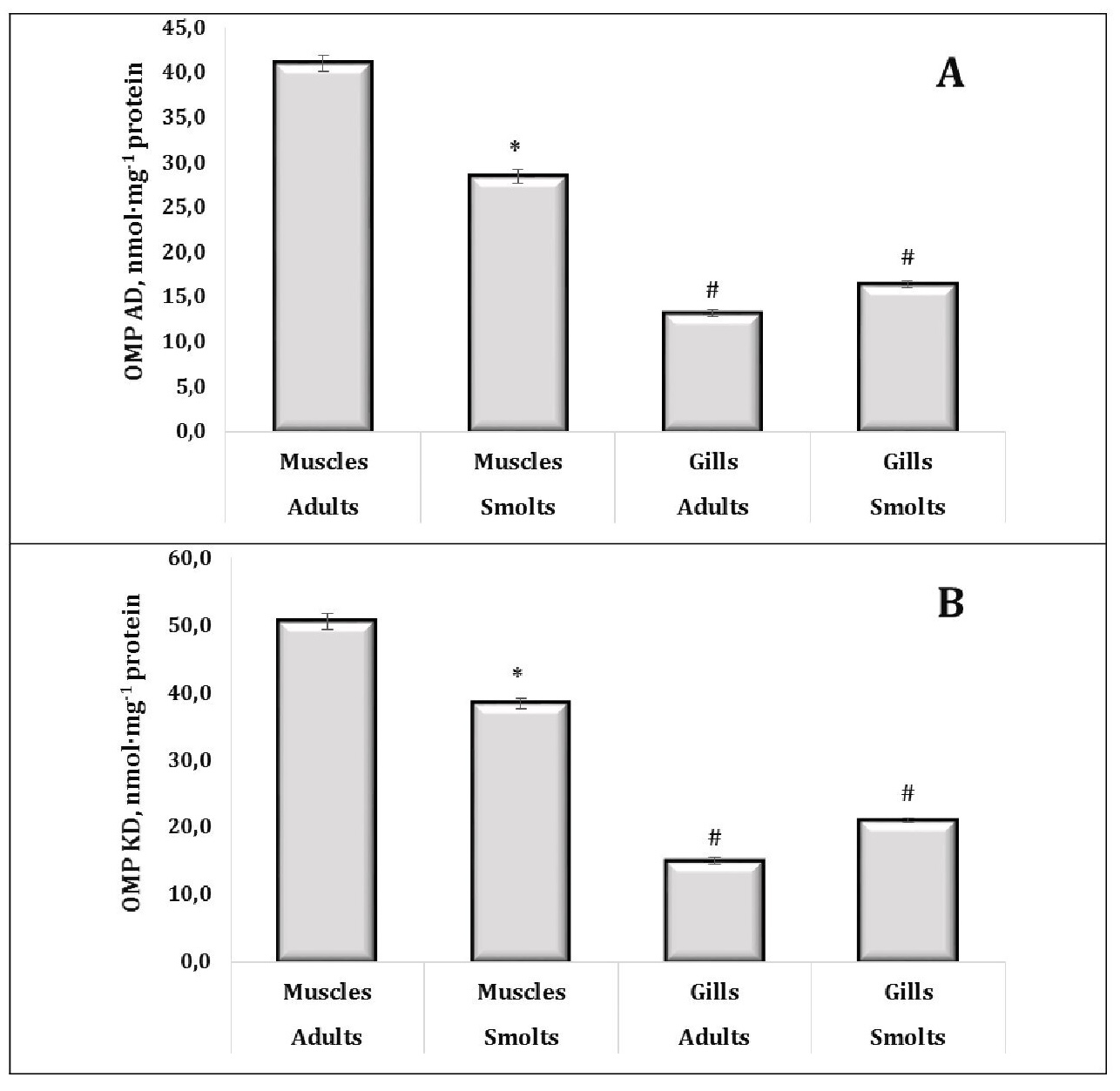
Fig. 7: Levels of aliphatic aldehyde dinitrophenylhydrazones (A, OMP AD, nmol·mg-1 protein) and aliphatic ketone dinitrophenylhydrazones level (B, OMP KD, nmol·mg-1 protein) in muscle and gill tissues of adult and juvenile sea trout (Salmo trutta L.) sampled from the Pomeranian region (northern Poland). * Changes are significant (p < 0.05) for one tissue type and different age groups; # Changes are significant (p < 0.05) for one age group and different tissue types. Results are expressed as mean ± S.E.M. (adults, muscles, n = 69; smolts, muscles, n = 128; adults, gills, n = 50; smolts, gills, n = 60). Differences between groups were analysed using two-way ANOVA followed by Bonferroni's post-hoc test.
Our study indicates that adult trout experience higher levels of oxidative stress compared to smolts, as evidenced by increased lipid peroxidation processes estimated by TBARS levels and the oxidative protein modification score, with gills being more susceptible to oxidative damage than muscle tissue. Although the TAS data were higher in muscle than in gills, they were not affected by the developmental stage, suggesting a stable antioxidant defence in muscle. These results highlight the greater susceptibility of adult trout to environmental contaminants and oxidative stress, particularly in the gills, and underline the cumulative effects of prolonged exposure to contaminants in aquatic environments.
The MANOVA statistical analysis also used sigma-restricted parameterisation, which facilitates interpretation by comparing factor levels relative to a reference category. This approach ensures that the statistical model used is identifiable and allows clear examination of differences between developmental stages and tissue types (muscle or gill) of trout. Decomposition of the effective hypotheses parameter further clarified the contribution of each factor analysed, with the developmental stage and tissue type showing strong main effects and their interaction providing additional, although less pronounced, variance explanation.
Thus, our results indicate that the developmental stage of trout has a significant effect on the dependent variables, suggesting physiological changes throughout the life cycle of the fish. The tissue type also plays a critical role, with clear differences in the response between these tissues. Furthermore, the significant interaction between the stage and tissue implies that the effect of the developmental stage varies depending on the tissue, highlighting the complexity of oxidative stress processes in fish at different life stages.
The next statistical analysis focused on evaluating the concentrations of selected chemical elements and oxidative stress markers in fish tissues at different developmental stages. The SS test for the full model versus SS for residuals compares the variability explained by the model with the unexplained variability, providing an assessment of model fit. A higher full model SS and a lower residual SS, together with a significant F statistic, indicate that the model effectively explains the variability in the data. The primary objective was to determine the extent to which these variables were influenced by the developmental stage, tissue type, and their interaction, providing insight into oxidative stress responses and potential toxicological effects (Table 1).
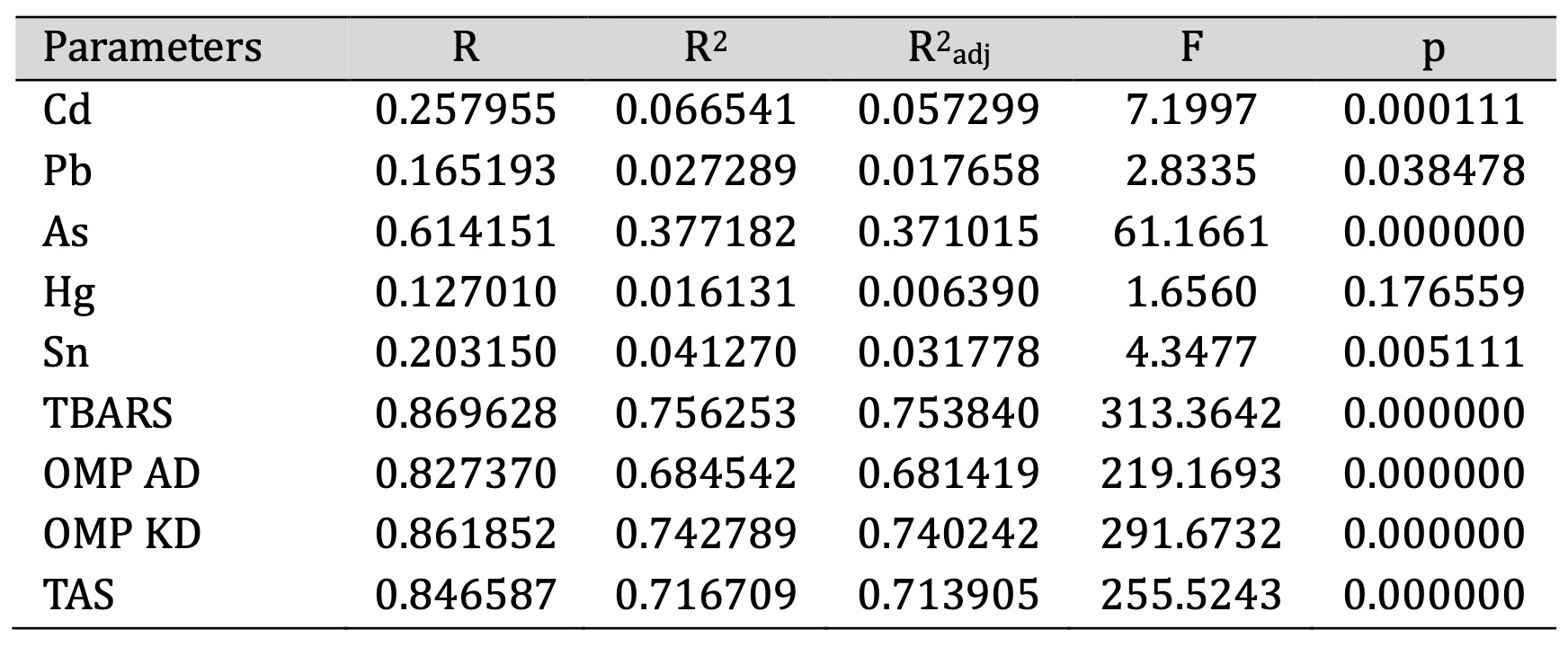
Table 1: SS test for the full model versus SS test for the MANOVA residuals for toxic metals and markers of oxidative stress in fish tissue
Cadmium showed a statistically significant relationship with an R² of 0.0665, indicating that 6.65% of the variance in the Cd concentration was explained by our model. The F value was 7.20 (p = 0.000), indicating strong evidence of a relationship. Lead showed a weaker association with an R² of 0.0273 and an F-value of 2.83 (p = 0.039), which was still statistically significant at the p = 0.05 level. Arsenic showed the strongest relationship among the metals analysed, with an R² of 0.3772, explaining 37.72% of the variance, and an F-statistic of 61.17 (p = 0.000). In contrast, mercury did not show a statistically significant relationship (R² = 0.0161, F = 1.66, p = 0.1766). Tin (Sn) showed a moderate association with an R² of 0.0413, an F-value of 4.35, and a p-value of 0.005, confirming a significant but relatively small effect (Table 1).
The model showed similar explanatory power for oxidatively modified proteins (OMP AD and OMP KD). OMP AD had an R² of 0.6845 (F = 219.17, p = 0.000), which means that 68.45% of the variance was attributable to the factors investigated. OMP KD showed an even stronger relationship with an R² of 0.7428 and an F-value of 291.67 (p = 0.000). These results indicate that both biomarkers of oxidative stress are significantly influenced by the independent variables, highlighting the role of oxidative damage in the tissues studied.
The analysis of the TAS level, which influences the total antioxidant defence status, showed an R² of 0.7167, indicating that 71.67% of the variance in the TAS value was explained by our statistical model. The F-value was 255.52 (p = 0.000), confirming the robustness of the relationship. These results emphasise that the factors investigated have a significant impact on the antioxidant capacity of fish tissues and further support the relevance of oxidative stress processes across developmental stages and tissue types.
Therefore, our analysis revealed a higher significance of oxidative stress markers (TBARS, OMP, TAS) compared to chemical elements, as indicated by the higher R values and the stronger F statistics. This suggests that oxidative stress responses were more strongly influenced by the independent variables, emphasising their greater relevance in the context of the study.
The regression analysis showed that the tissue factor had a significant negative effect on the Cd levels, as indicated by the t-value, which shows statistical significance. Furthermore, a strong and significant effect was observed in the interaction between the Stage and Tissue factors, which had a positive effect on the cadmium levels (p = 0.000). These results suggest that, while the Tissue factor may have had a negative effect on the Cd levels, the interaction between Stage and Tissue had a positive effect (Table 2).
Moving on to another element, the regression analysis showed that the Tissue variable had a positive effect on the Pb levels, with a significant relationship indicated by the coefficient and the t-value. The positive coefficient means that an increase in the Tissue variable is associated with an increase in the Pb levels. The statistical significance, with a p-value of less than 0.05, confirms that the relationship between Tissue and Pb is significant.
In the case of arsenic, the regression analysis shows that the Tissue variable had a significant negative effect on the As levels, demonstrating a strong and statistically significant negative relationship. Conversely, the Stage variable had a positive effect on the As levels, indicating a significant positive relationship. Both variables had a significant effect on the arsenic levels, with Tissue having a negative effect and Stage having a positive effect.
Similarly, to As, in the case of mercury, the Tissue variable had a positive effect on the Hg levels, with a statistically significant relationship indicated by the coefficient and the t-value. Specifically, the positive coefficient indicates that an increase in the Tissue variable corresponds to an increase in the Hg levels. The p-value of 0.041 confirms the statistical significance of this relationship (Table 2).
Next, the analysis of tin showed that the interaction between the Stage and Tissue factors had a significant negative effect on the Sn levels. The negative β-coefficient indicates that changes in both Stage and Tissue factors have a negative effect on the Sn levels. This relationship is statistically significant, confirming the influence of this interaction on the Sn levels (Table 2).
The regression analysis also showed that both Stage and Tissue had a significant negative effect on the TBARS levels, while the interaction between Stage and Tissue had a positive effect. Specifically, Stage had a strong negative relationship with TBARS, and higher Tissue values were associated with lower TBARS levels. On the other hand, the interaction between Stage and Tissue had a positive effect on TBARS levels, corresponding to the influence of chemical elements. All these relationships are statistically significant, confirming their influence on the TBARS levels. Notably, high β values were obtained for the other oxidative stress data (OMP AD, OMP KD, and TAS value), depending on the factors involved in the analysis (Table 2).
In summary, the regression analysis indicates that different factors (stage, tissue, and their interactions) significantly influence the levels of chemical elements, such as Cd, Pb, As, Hg, and Sn, as well as biomarkers of oxidative stress in trout. The results show different relationships between these variables, with some factors having distinct effects on the levels of these elements. Furthermore, the oxidative stress data also show significant relationships, with high β values indicating strong dependencies on the factors studied. The high statistical significance of all the relationships shown in Table 2 suggests that the factors studied play a crucial role in determining the concentration of these elements in fish in different environmental conditions. These results may reflect the influence of different environmental conditions, such as freshwater in rivers or saltwater in the sea, on trout. Overall, the analysis highlights the complex interactions and dependencies that influence elemental levels and oxidative stress and underlines the importance of considering multiple variables when examining the impact of environmental and biological data on fish organisms.
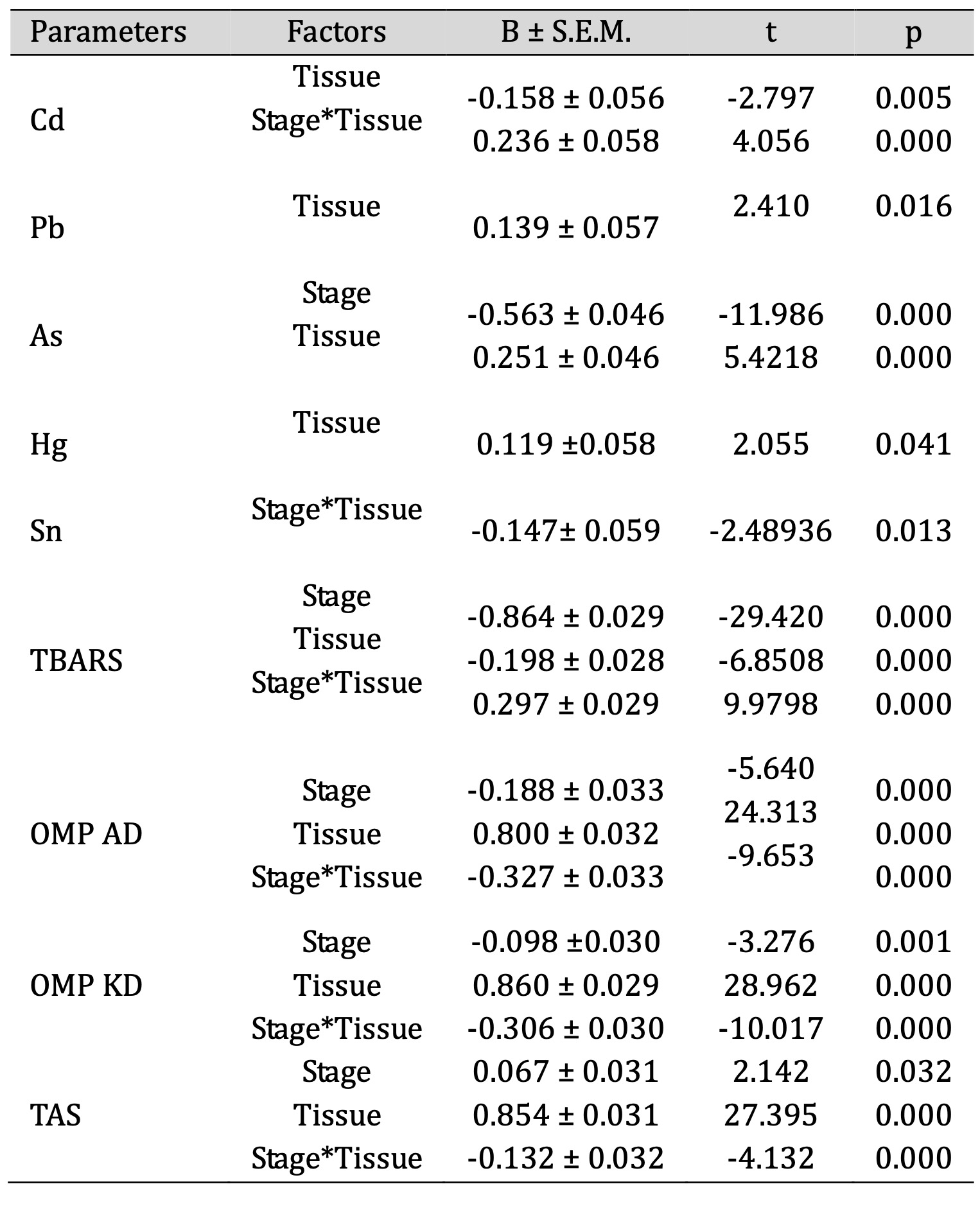
Table 2: Results of the regression analysis for the dependent variable (toxic metals: Cd, Pb, As, Hg, Sn) in trout, t values, and standardised regression coefficients (β ± S.E.M.) for the independent variables (Stage, Tissue, and Stage * Tissue)
Significant correlations between chemical elements and oxidative processes were also observed in the trout smolt muscle tissue (Table 3). The positive correlation between Pb and As suggests possible co-accumulation of these metals. In addition, the negative correlation between Pb and OMP KD indicates that higher Pb levels may be associated with reduced oxidative damage, possibly due to compensatory mechanisms in smolts. The almost perfect positive correlation between Hg and Sn suggests that these two metals co-accumulate in smolt muscle tissue, possibly reflecting similar bioaccumulation pathways. These results highlight the interactions between metal accumulation and oxidative damage in trout smolts and emphasise the complex relationships between environmental pollutants and physiological processes in early life stages.
Among the significant correlations observed in the adult trout gills (Table 3), the strongest correlation was found between Cd and Pb, indicating a positive relationship between the levels of accumulation of these two metals. In addition, the positive correlation between Cd and TBARS indicates that the increased Cd accumulation is associated with higher levels of oxidative stress. Regarding antioxidant status, As showed a negative correlation with TAS, suggesting that higher arsenic levels may reduce the ability of trout gills to combat oxidative stress. A strong negative correlation was also observed between Hg and TAS (r = –0.669, p = 0.000), suggesting that mercury accumulation severely impairs gill antioxidant defences. Finally, the positive correlation between TBARS and OMP AD reflects the link between lipid peroxidation and protein damage in response to oxidative stress in adult trout gills.
Based on our correlation analysis of chemical elements and oxidative stress indicators at both developmental stages (smolt and adult) and tissue types (muscle and gills), several key findings emerge. In adult trout, the accumulation of such metals as Cd, Pb, and As in the gills showed significant positive correlations, with Cd strongly correlated with Pb and As. In contrast, the trout smolts showed a different pattern of metal accumulation and oxidative stress. The Pb accumulation showed positive correlations with As, Sn, and TBARS and OMP AD oxidative stress markers, suggesting a complex interaction between these metals and their combined effect on oxidative damage. A remarkable correlation was observed between Hg and both TBARS and OMP AD, further highlighting the contribution of mercury to oxidative stress.
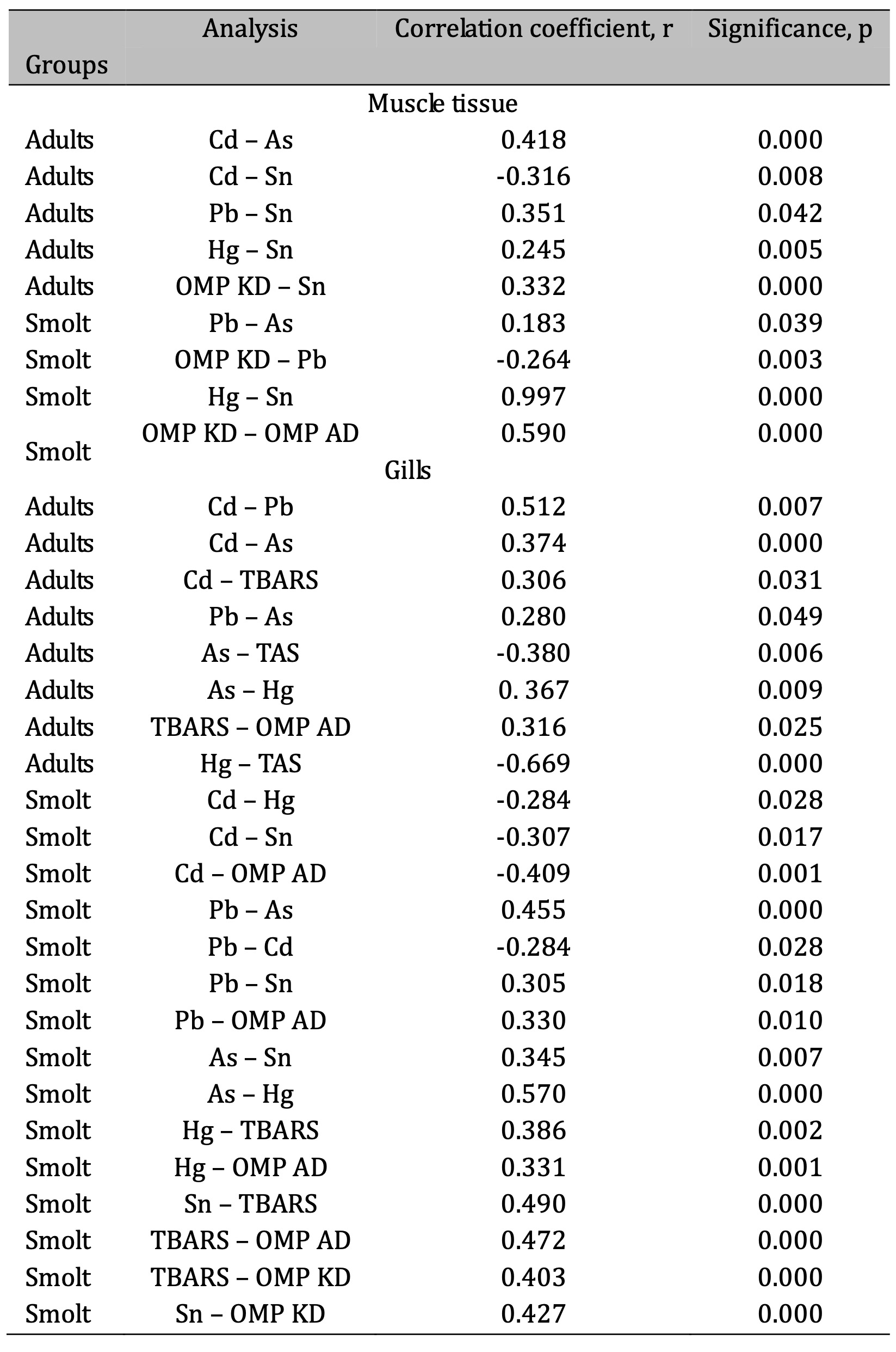
Table 3: Correlation analysis of toxic metal accumulation in trout muscle and gills at different developmental stages (adult vs. juvenile) based on r and p values
Discussion
The studies conducted aimed to analyse the interactions between oxidative stress parameters and toxic elements in muscle and gill tissues of smolt and adult trout. The results obtained highlight several important findings. Firstly, the study highlights distinct differences in the accumulation patterns of toxic elements, such as Cd, Pb, As, Hg, and Sn, in the muscle tissues and gills of sea trout (Salmo trutta L.), with notable variations observed between smolts and adults. In particular, the Cd levels were statistically higher in the gills of adult trout compared to muscle tissues, suggesting that the gills serve as a primary site for metal uptake in adults. Conversely, the smolt trout had higher total Cd levels, indicating age-related differences in metal uptake and detoxification processes. The adult muscle tissues showed a greater tendency to accumulate Pb and As, with statistically significant differences observed. Furthermore, arsenic showed a similar accumulation pattern in the adult gills, highlighting the dual role of these organs in element accumulation and exposure pathways.
Tin accumulation was also significantly higher in muscle tissues of adult trout compared to smolts, suggesting a cumulative effect over the lifetime of the trout. Taken together, these results highlight the critical role of age, tissue type and environmental exposure in shaping the bioaccumulation of chemical elements in migrating trout. The observed differences between gill and muscle tissues provide valuable insights into the physiological and ecological factors driving chemical element accumulation and highlight the importance of targeted monitoring and mitigation strategies to protect sensitive fish populations.
The phenomenon of cross-acclimation, as analysed in studies by McGeer et al . [47] and Hollis et al . [48], refers to the ability of an organism to adapt to one metal by influencing its response to and uptake of another metal. Prior exposure to one metal (e.g. copper) can alter the uptake and distribution of other metals (e.g. cadmium) in fish, with important implications for metal bioaccumulation and toxicity. McGeer et al . [47] demonstrated cross-acclimation where prior exposure to copper resulted in an increased capacity and affinity to accumulate cadmium in rainbow trout, while exposure to zinc had no similar effect. Copper-acclimated trout had a higher cadmium uptake capacity compared to unexposed controls, whereas cadmium-acclimated fish had a lower cadmium uptake rate. This study highlights the importance of considering past metal exposure when assessing toxicity and bioaccumulation, as acclimation can significantly influence an organism’s response to subsequent metal exposure [47]. Chowdhury et al . confirmed these findings by exposing rainbow trout to sublethal concentrations of waterborne and dietary cadmium, which induced a significant increase in metallothionein (MT) levels in various tissues [49]. The highest MT levels were observed in kidney and intestine tissues, suggesting competition between Cd and other metals (Zn, Cu) for binding sites on MT and non-MT proteins. This competition resulted in a marked decrease in Cd, Zn, Cu, and MT levels in the posterior intestine of acclimated fish following acute Cd-induced exposure, as shown by Chowdhury et al . [49]. This phenomenon is crucial for understanding how acclimation to a metal can alter the response of an organism to subsequent exposures, with implications for metal bioaccumulation and toxicity in aquatic organisms.
Our study showed that the accumulation of Cd, Pb, As, Hg, and Sn in the muscle tissue and gills of sea trout was particularly pronounced in adult specimens, as indicated by the statistically significant differences. Muscle tissue, being metabolically active and an important storage site, serves as a long-term reservoir for these metals. The persistent accumulation in muscle tissue reflects chronic exposure throughout the life cycle of trout, particularly in polluted environments, such as rivers and the Baltic Sea. For example, cadmium and mercury are known to bind strongly to proteins in muscle tissue, disrupting cellular metabolism and contributing to oxidative stress [7]. These findings are consistent with our statistical analysis, which showed a significant increase in the concentrations of these metals in the adult fish, suggesting that bioaccumulation increases with age and prolonged exposure to contaminated habitats.
The gills, a primary site of interaction with the aquatic environment [50, 51], also showed significant accumulation of chemical elements in adult trout, highlighting their role as a critical point of entry for contaminants. The gills are directly exposed to waterborne metals and act as a filtering organ, resulting in the deposition of such metals as lead and arsenic in their tissues [50, 51]. The higher accumulation observed in adult trout suggests that prolonged exposure to polluted waters during migration results in increased metal concentrations. These changes were statistically significant, indicating that gills are not only sensitive to acute exposure but also reflect chronic contamination levels. The combined effects of chemical element accumulation in muscle tissue and gills highlight the need for ongoing monitoring and conservation efforts to mitigate health risks to trout populations in polluted ecosystems.
Our previous study [34] showing that contaminant levels in Baltic Sea trout remain below internationally acceptable levels is crucial in this context. They provide evidence that this species can remain a relatively safe and nutritious food source despite environmental pressures. Such results are important for public health, as they reassure consumers about the safety of eating Baltic fish, while emphasising the need for continued monitoring and sustainable management of the Baltic Sea ecosystem. The study on the mineral composition of Baltic Sea trout and the levels of heavy metals has important implications not only for human health, but also for understanding the state of the Baltic Sea ecosystems. The results, which show that the consumption of sea trout provides substantial amounts of copper and magnesium while maintaining minimal levels of toxic heavy metals such as arsenic, lead, cadmium and mercury, indicate that the fish is of relatively high quality as a food source.
These results [34] have wider implications for the ecosystems of the Baltic Sea. As one of the most polluted seas in the world, the Baltic Sea is particularly vulnerable to the accumulation of toxic substances due to limited water exchange, intensive human activities and the high population density in its catchment area. The data confirming low concentrations of heavy metals in sea trout suggest that in some parts of the Baltic, ecological conditions remain stable enough to support healthy fish populations with low levels of bioaccumulated contaminants. These findings also highlight the delicate balance between exploiting marine resources and protecting the health of the environment. Effective mitigation strategies – such as reducing pollution at source, implementing stricter regulations and promoting international cooperation between countries bordering the Baltic Sea – are essential to ensure the long-term sustainability of both the ecosystem and the food resources it provides. Addressing the risk of pollution protects biodiversity and the health of communities that depend on the Baltic Sea, ensuring that it remains a viable habitat and valuable resource for future generations.
Telli-Karakoç and Barlas analysed the health of fish from selected aquacultures and observed rare abnormalities in such organs as the liver, spleen, intestines, and reproductive system [52]. However, these abnormalities did not significantly affect the overall health of the fish. Enzyme activity and protein concentrations varied mainly with age and season, rather than with farm location or environmental conditions, unless extreme or polluted. The concentrations of nickel (Ni), copper (Cu), zinc (Zn), arsenic (As), cadmium (Cd), lead (Pb), and mercury (Hg) did not exceed regulatory limits, indicating that fish in optimal farming conditions are of good quality and safe for human consumption [52].
A study by Meland et al . highlighted the long-term environmental impact of road runoff on aquatic ecosystems, similar to previous studies on the impact of metal exposure on fish health [53]. Traffic-related pollutants, including metals and polycyclic aromatic hydrocarbons (PAHs), were found in higher concentrations in tunnel wash water discharged into the Arungselva River, exceeding environmental quality standards [53]. These contaminants, which are thought to originate from tyre wear, brakes, and road salt, may have a negative impact on the fish population, as evidenced by the reduced growth of sea trout in downstream reaches, suggesting long-term ecological effects. PAHs and such elements as Al, Cd, Cr, Cu, Fe, and Pb were mainly associated with particles and colloids, while As, Ca, K, Mg, Mo, Ni, Sb, and Zn were more commonly found in their dissolved forms [53].
Our studies provide important insights into environmental factors influencing chemical element accumulation and oxidative stress in fish, particularly sea trout. Understanding the levels of such elements as Cd, Pb, As, Hg, and Sn is essential for assessing the health of aquatic ecosystems and the safety of seafood [54]. Toxic metal contamination is a major global concern, as it affects aquatic life and human health [54, 55]. By investigating these relationships, the study contributes to better environmental monitoring and the development of strategies to reduce water pollution [56].
Secondly, our results suggest that the increased oxidative stress observed in adult trout, as indicated by the increased TBARS levels and oxidative protein modifications, is likely to be related to the accumulation of chemical elements, such as Cd, Pb, As, Hg, and Sn. Gills, which are directly exposed to aquatic pollutants, showed greater oxidative damage than muscle tissue, consistent with the higher concentration of metals found in gills. While the muscle tissue had a higher TAS value, both the muscle and gill tissues of adult trout experienced significant oxidative stress, indicating the long-term effects of chemical element exposure. These findings highlight the detrimental effects of metal pollution on trout health, particularly in terms of oxidative damage, and emphasise the need to protect aquatic environments from ongoing pollution. It is also important to consider that other factors may contribute to the observed oxidative stress in adult trout, including such environmental variables as water temperature, oxygen levels, and the presence of other contaminants, e.g. plastics [57]. These factors can interact with toxic metals, potentially exacerbating their toxic effects and affecting the overall health of fish. In addition, such biological factors as trout age, sex, metabolic rate, and immune system responses may modulate the severity of oxidative damage [58]. Therefore, while metal accumulation is a key factor, it is crucial to consider the complex interplay of environmental and biological influences on the health of sea trout.
Kumar et al . have shown that such metals as cadmium, lead, mercury, and zinc significantly contribute to oxidative stress processes in sea trout, disrupting vital cellular functions and transcriptional profiling of genes [59]. Studies have also elucidated how toxic metals increase the production of reactive oxygen species (ROS) via Fenton-like reactions and mitochondrial dysfunction, while inhibiting key antioxidant enzymes, such as superoxide dismutase, catalase, and glutathione peroxidase, as demonstrated by Jomova et al . [60]. In addition, Srikanth et al . showed that depletion of non-enzymatic antioxidants, such as glutathione, weakens the ability of fish to counteract oxidative damage [61]. This imbalance leads to lipid peroxidation, protein oxidation, and DNA damage, causing structural and functional disruptions in cell membranes and proteins [62]. These findings highlight that oxidative stress impairs growth, immune function, and reproduction in sea trout, posing a significant risk to the health of the population.
This research is highly relevant in the context of current global environmental challenges [65]. The Baltic Sea is known for its relatively low water exchange with other oceans, leading to higher retention of pollutants in the ecosystem. The low water circulation combined with the shallow depth of a considerable part of the Baltic Sea means that pollutants can remain in the water for long periods, creating long-term exposure risks for marine life. For migrating trout, which are sensitive to changes in water quality and temperature, this prolonged exposure to pollutants can lead to severe physiological stress. This stress can be exacerbated by such factors as rising water temperatures, oxygen depletion, and eutrophication, i.e. conditions exacerbated by climate change and human activities [36]. Understanding the effects of different environmental conditions (freshwater versus saltwater) on the bioaccumulation of toxic elements is crucial, especially in the context of climate change, which is altering aquatic environments and influencing the behaviour of pollutants [11]. The results contribute to the field by addressing these issues and shedding light on the possible impact of different environmental factors on the health of aquatic species in different habitats.
Thirdly, the highest correlations observed in the adult trout gills highlight the significant interactions between metal accumulation and oxidative stress (Table 3). The strong positive correlations between Cd and Pb and between Cd and As suggest that these metals accumulate together in the gills, potentially increasing the overall toxic load. The positive correlation between Cd and TBARS supports the idea that metal accumulation is closely linked to increased oxidative stress, with lipid peroxidation serving as a marker of this process. In addition, the negative correlation between Hg and TAS highlights the detrimental effect of mercury on antioxidant status, further exacerbating oxidative damage in the gills. These results emphasise the complex relationships between metal accumulation and oxidative stress and highlight the potential threat posed by contaminants, particularly mercury, to the health of adult trout in contaminated environments. Significant correlations between chemical elements and oxidative processes in the gills and muscle tissues at the smolt and adult stages of sea trout are shown in Fig. 8.
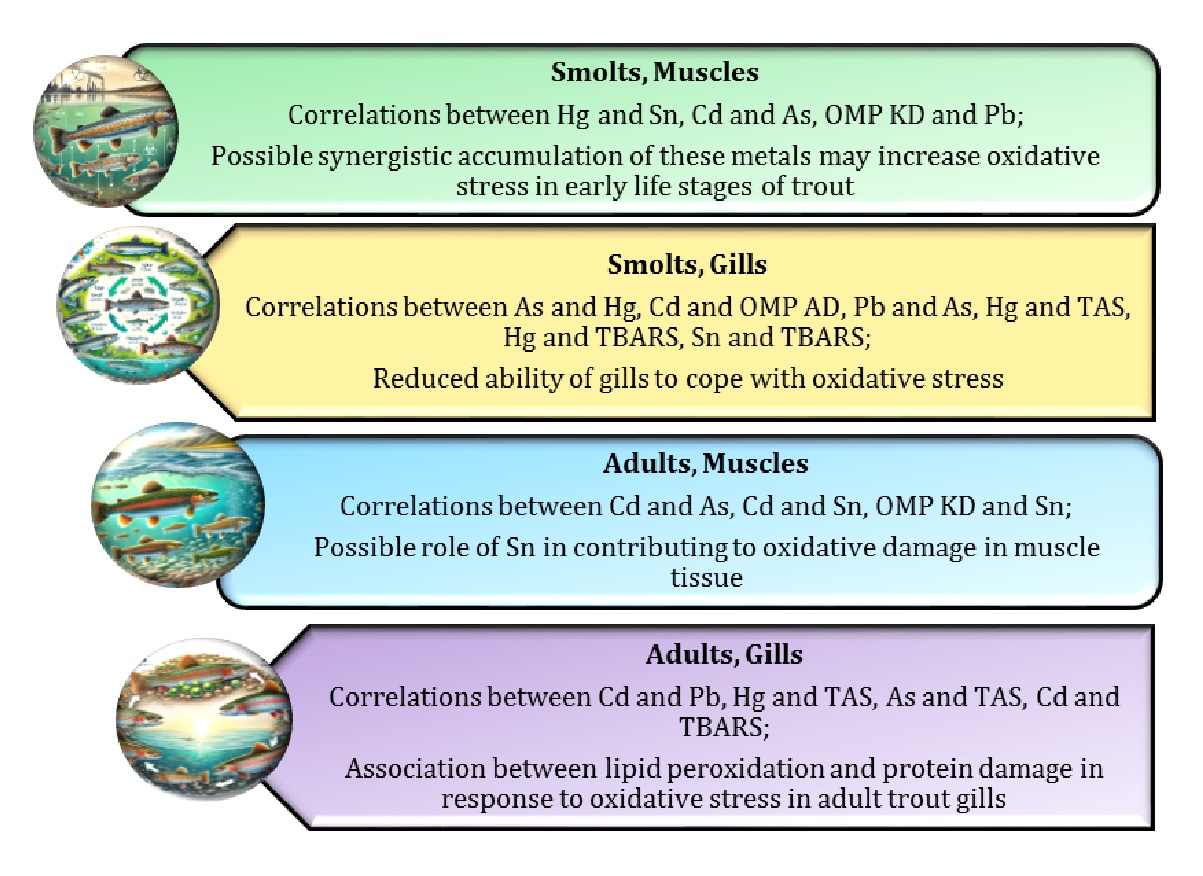
Fig. 8: Significant correlations between toxic metals and oxidative processes in gills and muscle tissues of smolt and adult sea trout.
Our results highlight the important role of tin in the biochemical processes in juvenile fish, although the existing literature on the effects of tin is sparse. Winship’s study showed that tin plays a complex role in aquatic environments and organisms, including fish [66]. Inorganic tin salts exhibit low toxicity due to poor absorption in the gastrointestinal tract and rapid excretion, with only a small fraction deposited in such tissues as lungs and bones. Conversely, organostannic compounds can be toxic, affecting fish through such mechanisms as mitochondrial dysfunction, membrane alterations, and neurotoxicity, mainly induced by trimethyltin and triethyltin compounds. These effects can disrupt key physiological processes, including oxidative phosphorylation and immunity, with potential implications for fish health and survival, as reported by Winship [66]. Although tin is not essential for fish or humans and no tin deficiency has been observed, certain compounds exhibit properties that could be exploited in medical applications, highlighting the need for careful risk assessment in aquatic systems, as demonstrated by Nagy et al . [67].
The novelty of our research lies in its comprehensive approach to studying the complex interactions between contaminant accumulation and biomarkers of oxidative stress in fish at different developmental stages. In addition, the unique environmental conditions of the Baltic Sea, including low oxygen levels in certain areas, may further complicate the health of migrating trout. Hypoxia, or low oxygen, is a common problem in the Baltic Sea, especially in deeper areas where water circulation is restricted [22]. Trout entering these areas may experience respiratory stress, reducing their ability to effectively process pollutants and potentially exacerbating the toxic effects of metals and other contaminants [5]. These combined stressors can significantly affect the survival and health of migratory fish species, making the Baltic Sea a particularly hazardous environment for such species as trout that migrate between freshwater and marine habitats. By integrating chemical element analysis with oxidative stress data and considering the effects of both freshwater and saltwater environments, our study provides a holistic understanding of the effect of pollution on aquatic life [55]. Our interdisciplinary approach linking environmental variables with biological markers of oxidative stress represents a novel contribution to ecological and environmental studies. Furthermore, the use of regression analysis to examine these interactions allows detailed exploration of the influence of specific factors on contaminant levels, providing valuable insights for future research and policy development.
Fourthly, our regression analysis indicates that several factors (developmental stage, tissue type and their interactions) significantly affect the levels of chemical elements such as Cd, Pb, As, Hg and Sn as well as oxidative stress biomarkers in trout, as shown in Table 2, underlining the critical role of these factors in determining the concentration of these elements in fish, particularly in response to environmental conditions. We would like to emphasise that the smolt gill data revealed several strong correlations that underline the complex relationships between metal accumulation and oxidative stress, as shown in Table 3. Sn was significantly correlated with both TBARS and OMP AD, highlighting its role in promoting oxidative damage in smolt gills. The strong positive correlations between oxidative stress indicators such as TBARS and OMP AD further support the concept of integrated oxidative damage in response to metal accumulation. Fig. 9 summarises the key findings of our study and illustrates the impact of chemical element contamination on trout health, with a focus on oxidative damage.

Fig. 9: The figure summarises the key findings of our study and illustrates the impact of chemical element contamination on trout health, with a focus on oxidative damage.
A limitation of the present study is the omission of sex as a biological variable. Although sex differences in metal accumulation have been reported in fish, it was not possible to reliably determine the sex of all individuals, particularly at the smolt stage where there is no external sexual dimorphism. Including only individuals of known sex would have significantly reduced the sample size and hence the statistical power of the analysis. Future investigations should aim to include sex as a factor to better understand its potential influence on metal bioaccumulation patterns.
Future research on the effects of chemical elements on oxidative stress in sea trout and other aquatic species has considerable potential to advance our understanding of environmental toxicity and its biological effects. Researchers could carry out long-term multi-generational studies to determine the impact of chronic exposure on genetic and epigenetic adaptations. In addition, studying the combined effects of chemical elements with other stressors, such as climate change or habitat degradation, would provide a more comprehensive view of ecosystem health [68, 69]. Future work should also focus on identifying biomarkers for early detection of oxidative stress and developing strategies to mitigate metal toxicity, such as bioremediation techniques or dietary supplementation with antioxidants. These studies are essential to inform conservation efforts, regulatory policies, and sustainable management practices for aquatic ecosystems.
Broader implications
In the study by Tkachenko et al. [34] analysed the mineral composition and heavy metal concentrations in the muscles of Baltic Sea trout to assess their suitability for human consumption. The results showed that the concentrations of the elements in the fish were below the maximum permissible levels set by international food safety standards, confirming no significant health risk associated with their consumption. Baltic Sea trout was found to be a particularly rich source of copper and magnesium in the diets of both children and adults, although levels of other minerals were less significant. In addition, fish consumption was not associated with significant exposure to heavy metals such as arsenic, lead, cadmium or mercury. This suggests that Baltic Sea trout can be a safe and valuable part of the diet, especially in providing essential minerals. The study highlights the potential of Baltic Sea trout as a beneficial dietary option due to its ability to provide key minerals such as copper and magnesium, while ensuring safety from toxic metal exposure. This is particularly important for the development of dietary strategies that improve mineral intake without compromising health standards [34].
Chandel et al . [31] showed that both natural and anthropogenic arsenic forms pose a significant threat to aquatic organisms, with bioaccumulation in fish potentially affecting higher trophic levels, including humans [31, 65]. Continued exposure to arsenic in aquatic environments underscores the urgent need for sustainable practices and policies to mitigate contamination and protect both aquatic ecosystems and human health [28]. Arsenic interferes with enzymes involved in cellular energy production by binding to thiol groups, potentially affecting the physiological functions of fish, including their nervous systems. This interference may lead to disruptions in neural pathways, potentially resulting in neurodegenerative changes, such as an altered protein composition in the cytoskeleton and hyperphosphorylation of cytoskeletal proteins. Such disruptions in the cytoskeletal structure can impair cellular integrity and function, affecting fish behaviour, sensory responses, and overall health [63].
Polak-Juszczak and Szlinder-Richert (2021) studied the impact of chemical weapons dumped in seas and oceans after World War II, including the Baltic Sea, on marine pollution. Sunken containers released toxic substances, such as arsenic, into the environment. The study examined total arsenic, inorganic arsenic (III + V), and organic arsenic compounds in the muscle tissue of cod, herring, sprat, and flounder. The authors reported that sprat had the highest total arsenic content, while cod had the lowest. The risk to consumer health, based on the estimated daily intake and carcinogenic risk, showed no significant health threat, with carcinogenic risk values within the acceptable range [64].
Thus, studies have demonstrated that Baltic Sea fish, including trout, generally contain essential minerals like copper and magnesium while maintaining low levels of toxic metals such as arsenic, cadmium, lead, and mercury, with concentrations typically below international safety limits. Nevertheless, historical pollution, e.g. the release of arsenic from sunken chemical weapons, in conjunction with ongoing environmental contamination, underscores the necessity for sustained monitoring due to the potential for bioaccumulation and its repercussions on aquatic organisms and ecosystem health.
Conclusion
Our results show significant tissue and age-related patterns of chemical element accumulation in sea trout. In adult trout, higher levels of Pb, As, Hg, and Sn were found in muscle tissues, whereas Cd accumulated predominantly in the gills, especially in smolts. These results emphasise the influence of age and tissue type on chemical element bioaccumulation. Our data suggest that the increased oxidative stress observed in adult trout, as indicated by increased lipid peroxidation (TBARS) and oxidative protein modifications, is likely to be driven by the accumulation of such chemical elements as Cd, Pb, As, Hg, and Sn. Gills directly exposed to waterborne contaminants showed more pronounced oxidative damage than muscle tissue, which correlated with higher chemical element concentrations found in the gills. Although the muscle tissue had a higher total antioxidant capacity, both muscle and gill tissues of adult trout experienced significant oxidative stress, reflecting the long-term effects of chemical element exposure. These findings highlight the detrimental effects of chemical pollutants on trout health, particularly in terms of oxidative damage, and emphasise the importance of protecting aquatic ecosystems from ongoing contamination.
The correlation and regression analysis of chemical elements and oxidative stress markers across the different developmental stages (smolt and adult) and tissue types (muscle and gills) revealed several key patterns. In the adult trout, significant positive correlations were observed between the accumulation of such elements as Cd, Pb, and As in the gills, with Cd showing a strong relationship with both Pb and As. Conversely, the trout smolts showed a distinct pattern of chemical element accumulation and oxidative stress. In particular, the Pb accumulation was positively correlated with As, Sn, and oxidative stress markers, such as TBARS and OMP AD, suggesting a complex interaction between these metals and their collective effect on oxidative damage. In addition, a significant correlation was found between Hg and both TBARS and OMP AD, highlighting the contribution of mercury to oxidative stress in trout smolts. These results provide valuable insights into the complex interactions between chemical element accumulation and oxidative stress in sea trout and highlight the need for comprehensive strategies to mitigate the effects of environmental pollutants on aquatic species.
Acknowledgements
Funding
This research received no specific grant from any funding agency, commercial or not-for-profit sectors.
Author contributions
N.K., H.T., P.K. – conceived the concept of the review; N.K., H.T. – developed the search strategy; N.K., H.T., P.K. – coordinated data selection, extraction, analysis, and interpretation; N.K., H.T. – critically reviewed the manuscript; N.K., H.T., P.K. – drafted the final manuscript.
Disclosure Statement
The authors have no competing interests to declare.
References
| 1 | Climate Change in the Baltic Sea. 2021 Fact Sheet. Baltic Sea Environment Proceedings n°180 HELCOM/Baltic Earth, 2021 ISSN: 0357-2994.
|
| 2 | Acioly, T.M.D.S.; da Silva, M.F.; Barbosa, L.A.; Iannacone, J.; Viana, D.C. Levels of Potentially Toxic and Essential Elements in Water and Estimation of Human Health Risks in a River Located at the Interface of Brazilian Savanna and Amazon Biomes (Tocantins River). Toxics 2024;12(7), 444
https://doi.org/10.3390/toxics12070444 |
| 3 | Pandelova, M.; Henkelmann, B.; Roots, O.; Simm, M.; Järv, L.; Benfenati, E.; Schramm, K.W. Levels of PCDD/F and dioxin-like PCB in Baltic fish of different age and gender. Chemosphere 2008, 71(2), 369-378
https://doi.org/10.1016/j.chemosphere.2007.08.050 |
| 4 | Kwasigroch, U.; Bełdowska, M.; Jędruch, A.; Łukawska-Matuszewska, K. Distribution and bioavailability of mercury in the surface sediments of the Baltic Sea. Environ. Sci. Pollut. Res. Int. 2021, 28(27), 35690-35708
https://doi.org/10.1007/s11356-021-13023-4 |
| 5 | Mackenzie, B.R.; Almesjö, L.; Hansson, S. Fish, fishing, and pollutant reduction in the Baltic Sea. Environ. Sci. Technol. 2004, 38(7), 1970-1976
https://doi.org/10.1021/es034297n |
| 6 | Cordeli, A.N.; Oprea, L.; Crețu, M.; Dediu, L.; Coadă, M.T.; Mînzală, D.-N. Bioaccumulation of Metals in Some Fish Species from the Romanian Danube River: A Review. Fishes 2023, 8(8), 387.
https://doi.org/10.3390/fishes8080387 |
| 7 | Song, C.; Sun, C.; Liu, B.; Xu, P. Oxidative Stress in Aquatic Organisms. Antioxidants (Basel) 2023, 12(6), 1223
https://doi.org/10.3390/antiox12061223 |
| 8 | Struciński, P.; Piskorska-Pliszczyńska, J.; Maszewski, S.; Góralczyk, K.; Warenik-Bany, M.; Mikołajczyk, S.; Czaja, K.; Hernik, A.; Ludwicki, J.K. PCDD/Fs and DL-PCBs intake from fish caught in Polish fishing grounds in the Baltic Sea - characterizing the risk for consumers. Environ. Int. 2013, 56, 32-41
https://doi.org/10.1016/j.envint.2013.03.002 |
| 9 | Duval, E.; Skaala, Ø.; Quintela, M.; Dahle, G.; Delaval, A.; Wennevik, V.; Glover, K.A.; Hansen, M.M. Long-term monitoring of a brown trout (Salmo trutta) population reveals kin-associated migration patterns and contributions by resident trout to the anadromous run. BMC Ecol. Evol. 2021, 21(1), 143
https://doi.org/10.1186/s12862-021-01876-9 |
| 10 | Strøm, J.F.; Jensen, J.L.A.; Nikolopoulos, A.; Nordli, E.; Bjørn, P.A.; Bøhn, T. Sea trout Salmo trutta in the subarctic: home-bound but large variation in migratory behaviour between and within populations. J. Fish Biol. 2021, 99(4), 1280-1291
https://doi.org/10.1111/jfb.14832 |
| 11 | Jonsson, B.; Jonsson, N. Migratory timing, marine survival and growth of anadromous brown trout Salmo trutta in the River Imsa, Norway. J. Fish Biol. 2009, 74(3), 621-638
https://doi.org/10.1111/j.1095-8649.2008.02152.x |
| 12 | Lynch, A.J.; Cooke, S.J.; Deines, A.M.; Bower, S.D.; Bunnell, D.B.; Cowx, I.G.; Nguyen, V.M.; Nohner, J.; Phouthavong, K.; Riley, B.; Rogers, M.W.; Taylor, W.W.; Woelmer, W.; Youn, S.J.; Douglas B.T. Jr. The social, economic, and environmental importance of inland fish and fisheries. Environ. Rev. 2016, 24(2), 115-121
https://doi.org/10.1139/er-2015-0064 |
| 13 | Harvey, A.C.; Glover, K.A.; Wennevik, V.; Skaala, Ø. Atlantic salmon and sea trout display synchronised smolt migration relative to linked environmental cues. Sci. Rep. 2020, 10(1), 3529
https://doi.org/10.1038/s41598-020-60588-0 |
| 14 | Pelletier, M.C.; Ebersole, J.; Mulvaney, K.; Rashleigh, B.; Gutierrez, M.N.; Chintala, M.; Kuhn, A.; Molina, M.; Bagley, M.; Lane, C. Resilience of aquatic systems: Review and management implications. Aquat. Sci. 2020, 82(2), 1-44
https://doi.org/10.1007/s00027-020-00717-z |
| 15 | Piper, A.T.; Rosewarne, P.J.; Bekkevold, D.; Grey, J.; Nye, A.; Wright, R.M. Migration patterns, habitat use and genetic origins of sea trout (Salmo trutta) in Norfolk chalk streams: implications for management of a mixed stock fishery. Aquat. Sci. 2025, 87, 7
https://doi.org/10.1007/s00027-024-01135-1 |
| 16 | Kurhaluk, N. Formation of an antioxidant profile in the sea trout (Salmo trutta m. trutta L.) from the Slupia River. Zoology (Jena) 2019, 133, 54-65
https://doi.org/10.1016/j.zool.2019.02.002 |
| 17 | HELCOM, 2011 Salmon and Sea Trout Populations and Rivers in the Baltic Sea - HELCOM assessment of salmon (Salmo salar) and sea trout (Salmo trutta) populations and habitats in rivers flowing to the Baltic Sea. Balt. Sea Environ. Proc. No. 126A. ISSN 0357-2994.
|
| 18 | Hoar, W.S. Smolt transformation: Evolution, behavior, and physiology. J. Fish. Board Can. 1976, 33, 1233-1252
https://doi.org/10.1139/f76-158 |
| 19 | Merola, C.; Bisegna, A.; Angelozzi, G.; Conte, A.; Abete, M.C.; Stella, C.; Pederiva, S.; Faggio, C.; Riganelli, N.; Perugini, M. Study of Heavy Metals Pollution and Vitellogenin Levels in Brown Trout (Salmo trutta trutta) Wild Fish Populations. Appl. Sci. 2021, 11(11), 4965
https://doi.org/10.3390/app11114965 |
| 20 | Angioni, A.; Corrias, F.; Alessandro, A.; Sabatini, A.; Palmas, F.; Lai, C.; Russo, M. Heavy metal and metalloid accumulation in wild brown trout (Salmo trutta L., 1758 complex, Osteichthyes: Salmonidae) from a mountain stream in Sardinia by ICP-OES. Environ. Monit. Assess. 2021, 193(7), 448
https://doi.org/10.1007/s10661-021-09204-w |
| 21 | Kurhaluk, N.; Tkachenko, H. Antioxidants, lysosomes and elements status during the life cycle of sea trout Salmo trutta m. trutta L. Sci. Rep. 2021, 11(1), 5545
https://doi.org/10.1038/s41598-021-85127-3 |
| 22 | Andersson, M.; Jonsson, B.; Calles, O.; Greenberg, L. Assessing Movements between Freshwater and Saltwater by Brown Trout (Salmo trutta L.) Based on Otolith Microchemistry. Animals (Basel) 2024, 14(14), 2116
https://doi.org/10.3390/ani14142116 |
| 23 | Birnie-Gauvin, K.; Thorstad, E.; Aarestrup, K. Overlooked aspects of the Salmo salar and Salmo trutta lifecycles. Rev. Fish Biol. Fish. 2019, 29, 749-766
https://doi.org/10.1007/s11160-019-09575-x |
| 24 | Bhat, R.A.; Bakhshalizadeh, S.; Guerrera, M.C.; Kesbiç, O.S.; Fazio, F. Toxic effect of heavy metals on ovarian deformities, apoptotic changes, oxidative stress, and steroid hormones in rainbow trout. J. Trace Elem. Med. Biol. 2023, 75, 127106
https://doi.org/10.1016/j.jtemb.2022.127106 |
| 25 | Farmen, E.; Harman, C.; Hylland, K.; Tollefsen, K.E. Produced water extracts from North Sea oil production platforms result in cellular oxidative stress in a rainbow trout in vitro bioassay. Mar. Pollut. Bull. 2010, 60(7), 1092-1098
https://doi.org/10.1016/j.marpolbul.2010.01.015 |
| 26 | Morro, B.; Broughton, R.; Balseiro, P.; Handeland, S.O.; Mackenzie, S.; Doherty, M.K.; Whitfield, P.D.; Shimizu, M.; Gorissen, M.; Sveier, H.; Albalat, A. Endoplasmic reticulum stress as a key mechanism in stunted growth of seawater rainbow trout (Oncorhynchus mykiss). BMC Genomics 2021, 22(1), 824
https://doi.org/10.1186/s12864-021-08153-5 |
| 27 | Saç, H.; Yeltekin, A.Ç. Investigation of oxidative stress status and apoptotic markers of juvenile trout exposed to arsenic toxicity. Toxicol. Res. (Camb.) 2023, 12(4), 608-614
https://doi.org/10.1093/toxres/tfad049 |
| 28 | Thomas, D.J.; Styblo, M.; Lin, S. The cellular metabolism and systemic toxicity of arsenic. Toxicol. Appl. Pharmacol. 2001, 176(2), 127-144
https://doi.org/10.1006/taap.2001.9258 |
| 29 | Ngo, H.T.T.; Nguyen, T.D.; Nguyen, T.T.H.; Le, T.T.; Nguyen, D.Q. Adverse Effects of Toxic Metal Pollution in Rivers on the Physiological Health of Fish. Toxics 2022, 10(9), 528
https://doi.org/10.3390/toxics10090528 |
| 30 | Jamil Emon, F.; Rohani, M.F.; Sumaiya, N.; Tuj Jannat, M.F.; Akter, Y.; Shahjahan, M.; Abdul Kari, Z.; Tahiluddin, A.B.; Goh, K.W. Bioaccumulation and Bioremediation of Heavy Metals in Fishes - A Review. Toxics 2023, 11(6), 510
https://doi.org/10.3390/toxics11060510 |
| 31 | Chandel, M.; Sharma, A.K.; Thakur, K.; Sharma, D.; Brar, B.; Mahajan, D.; Kumari, H.; Pankaj, P.P.; Kumar, R. Poison in the water: Arsenic's silent assault on fish health. J. Appl. Toxicol. 2024, 44(9), 1282-1301
https://doi.org/10.1002/jat.4581 |
| 32 | Hansen, B.H.; Rømma, S.; Garmo, Ø.A.; Olsvik, P.A.; Andersen, R.A. Antioxidative stress proteins and their gene expression in brown trout (Salmo trutta) from three rivers with different heavy metal levels. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 143(3), 263-274
https://doi.org/10.1016/j.cbpc.2006.02.010 |
| 33 | Durrant, C.J.; Stevens, J.R.; Hogstrand, C.; Bury, N.R. The effect of metal pollution on the population genetic structure of brown trout (Salmo trutta L.) residing in the River Hayle, Cornwall, UK. Environ. Pollut. 2011, 159(12), 3595-3603
https://doi.org/10.1016/j.envpol.2011.08.005 |
| 34 | Tkachenko H., Kasiyan O., Kamiński P., Kurhaluk N. Assesing the human health risk of Baltic Sea trout (Salmo trutta L) consumption. Fisheries & Aquatic Life 2022, 30, 27-43
https://doi.org/10.2478/aopf-2022-0003 |
| 35 | Simmons, O.M.; Britton, J.R.; Gillingham, P.K.; Nevoux, M.; Riley, W.D.; Rivot, E.; Gregory, S.D. Predicting how environmental conditions and smolt body length when entering the marine environment impact individual Atlantic salmon Salmo salar adult return rates. J. Fish Biol. 2022, 101(2), 378-388
https://doi.org/10.1111/jfb.14946 |
| 36 | Elliott, J.M.; Elliott, J.A. Temperature requirements of Atlantic salmon Salmo salar, brown trout Salmo trutta and Arctic charr Salvelinus alpinus: predicting the effects of climate change. J. Fish Biol. 2010, 77(8), 1793-1817
https://doi.org/10.1111/j.1095-8649.2010.02762.x |
| 37 | Obolewski, K. Use of Macroinvertebrates as Bioindicators for the Assessment of Surface Water Quality in Urban Areas: A Case Study. Ochrona Środowiska, 2010, 32(2), 35-42.
|
| 38 | Pratten, D.J.; Shearer W.M. Sea Trout of the North Esk. Aquaculture Res. 1983,
https://doi.org/10.1111/j.1365-2109.1983.tb00056.x |
| 39 | Pedersen, S.; Christiansen, R.; Glüsing, H. Comparison of Survival, Migration and Growth in Wild, Offspring from Wild (F1) and Domesticated Sea-Run Trout (Salmo trutta L.) In: Harris G. and Milner N., editor. Sea trout: Biology, Conservation, and Management. Oxford, Blackwell Publishing; 2007 pp. 377-388.
https://doi.org/10.1002/9780470996027.ch26 |
| 40 | Pemberton, R. Sea trout in North Argyll Sea lochs, population, distribution and movements. J. Fish Biol. 1976, 9, 157-179
https://doi.org/10.1111/j.1095-8649.1976.tb04670.x |
| 41 | Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302-310
https://doi.org/10.1016/S0076-6879(78)52032-6 |
| 42 | Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248-254
https://doi.org/10.1016/0003-2697(76)90527-3 |
| 43 | Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol. 1990, 186, 464-478
https://doi.org/10.1016/0076-6879(90)86141-H |
| 44 | Reznick, A.Z.; Packer, L. Oxidative damage to proteins: spectrophotometric method for carbonyl assay. Methods Enzymol. 1994, 233, 357-363
https://doi.org/10.1016/S0076-6879(94)33041-7 |
| 45 | Miller, N.J.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. (Lond.) 1993, 84(4), 407-412
https://doi.org/10.1042/cs0840407 |
| 46 | Zar, J.H. Biostatistical Analysis. 4th ed., Prentice-Hall Inc., Englewood Cliffs, New Jersey, 1999.
|
| 47 | McGeer, J.C.; Nadella, S.; Alsop, D.H.; Hollis, L.; Taylor, L.N.; McDonald, D.G.; Wood, C.M. Influence of acclimation and cross-acclimation of metals on acute Cd toxicity and Cd uptake and distribution in rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2007, 84(2), 190-197
https://doi.org/10.1016/j.aquatox.2007.03.023 |
| 48 | Hollis, L.; Hogstrand, C.; Wood, C.M. Tissue-specific cadmium accumulation, metallothionein induction, and tissue zinc and copper levels during chronic sublethal cadmium exposure in juvenile rainbow trout. Arch. Environ. Contam. Toxicol. 2001, 41(4), 468-474
https://doi.org/10.1007/s002440010273 |
| 49 | Chowdhury, M.J.; Baldisserotto, B.; Wood, C.M. Tissue-specific cadmium and metallothionein levels in rainbow trout chronically acclimated to waterborne or dietary cadmium. Arch. Environ. Contam. Toxicol. 2005, 48(3), 381-390
https://doi.org/10.1007/s00244-004-0068-2 |
| 50 | Evans, D.H. The fish gill: site of action and model for toxic effects of environmental pollutants. Environ. Health Perspect. 1987, 71, 47-58
https://doi.org/10.1289/ehp.877147 |
| 51 | Evans, D.H.; Piermarini, P.M.; Choe, K.P. The multifunctional fish gill: dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 2005, 85(1), 97-177
https://doi.org/10.1152/physrev.00050.2003 |
| 52 | Telli-Karakoç, F.; Barlas, N. Determination of the healthiness of aquaculture fish by enzymes and histopathological methods. Mar. Pollut. Bull. 2019, 149, 110535
https://doi.org/10.1016/j.marpolbul.2019.110535 |
| 53 | Meland, S.; Borgstrøm, R.; Heier, L.S.; Rosseland, B.O.; Lindholm, O.; Salbu, B. Chemical and ecological effects of contaminated tunnel wash water runoff to a small Norwegian stream. Sci. Total Environ. 2010, 408(19), 4107-4117
https://doi.org/10.1016/j.scitotenv.2010.05.034 |
| 54 | Saravanan, P.; Saravanan, V.; Rajeshkannan, R.; Arnica, G.; Rajasimman, M.; Baskar, G.; Pugazhendhi, A. Comprehensive review on toxic heavy metals in the aquatic system: sources, identification, treatment strategies, and health risk assessment. Environ. Res. 2024, 258, 119440
https://doi.org/10.1016/j.envres.2024.119440 |
| 55 | Hama Aziz, K.H.; Mustafa, F.S.; Omer, K.M.; Hama, S.; Hamarawf, R.F.; Rahman, K.O. Heavy metal pollution in the aquatic environment: efficient and low-cost removal approaches to eliminate their toxicity: a review. RSC Adv. 2023, 13(26), 17595-17610
https://doi.org/10.1039/D3RA00723E |
| 56 | Singh, V.; Singh, N.; Rai, S.N.; Kumar, A.; Singh, A.K.; Singh, M.P.; Sahoo, A.; Shekhar, S.; Vamanu, E.; Mishra, V. Heavy Metal Contamination in the Aquatic Ecosystem: Toxicity and Its Remediation Using Eco-Friendly Approaches. Toxics 2023, 11(2), 147
https://doi.org/10.3390/toxics11020147 |
| 57 | Subaramaniyam, U.; Allimuthu, R.S.; Vappu, S.; Ramalingam, D.; Balan, R.; Paital, B.; Panda, N.; Rath, P.K.; Ramalingam, N.; Sahoo, D.K. Effects of microplastics, pesticides and nano-materials on fish health, oxidative stress and antioxidant defense mechanism. Front. Physiol. 2023, 14, 1217666
https://doi.org/10.3389/fphys.2023.1217666 |
| 58 | Canosa, L.F.; Bertucci, J.I. The effect of environmental stressors on growth in fish and its endocrine control. Front. Endocrinol. (Lausanne) 2023, 14, 1109461
https://doi.org/10.3389/fendo.2023.1109461 |
| 59 | Kumar, M.; Singh, S.; Jain, A.; Yadav, S.; Dubey, A.; Trivedi, S.P. A review on heavy metal-induced toxicity in fishes: Bioaccumulation, antioxidant defense system, histopathological manifestations, and transcriptional profiling of genes. J. Trace Elem. Med. Biol. 2024, 83, 127377
https://doi.org/10.1016/j.jtemb.2023.127377 |
| 60 | Jomova, K.; Raptova, R.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: chronic diseases and aging. Arch. Toxicol. 2023, 97(10), 2499-2574
https://doi.org/10.1007/s00204-023-03562-9 |
| 61 | Srikanth, K.; Pereira, E.; Duarte, A.C.; Ahmad, I. Glutathione and its dependent enzymes' modulatory responses to toxic metals and metalloids in fish - a review. Environ. Sci. Pollut. Res. Int. 2013, 20(4), 2133-2149
https://doi.org/10.1007/s11356-012-1459-y |
| 62 | Shahjahan, M.; Taslima, K.; Rahman, M.S.; Al-Emran, M.; Alam, S.I.; Faggio, C. Effects of heavy metals on fish physiology - A review. Chemosphere 2022, 300, 134519
https://doi.org/10.1016/j.chemosphere.2022.134519 |
| 63 | Vahidnia, A.; van der Voet, G.B.; de Wolff, F.A. Arsenic neurotoxicity - a review. Hum. Exp. Toxicol. 2007, 26(10), 823-832
https://doi.org/10.1177/0960327107084539 |
| 64 | Polak-Juszczak, L.; Szlinder Richert, J. Arsenic speciation in fish from Baltic Sea close to chemical munitions dumpsites. Chemosphere 2021, 284, 131326
https://doi.org/10.1016/j.chemosphere.2021.131326 |
| 65 | Valskienė, R.; Baršienė, J.; Butrimavičienė, L.; Grygiel, W.; Stunžėnas, V.; Jokšas, K.; Stankevičiūtė, M. Environmental genotoxicity and cytotoxicity levels in herring (Clupea harengus), flounder (Platichthys flesus) and cod (Gadus morhua) inhabiting the Gdansk Basin of the Baltic Sea. Mar. Pollut. Bull. 2018, 133, 65-76
https://doi.org/10.1016/j.marpolbul.2018.05.023 |
| 66 | Winship, K.A. Toxicity of tin and its compounds. Adverse Drug React. Acute Poisoning Rev. 1988, 7(1), 19-38.
|
| 67 | Nagy, L.; Szorcsik, A.; Kovács, K. Az ón és ónorganikus vegyületek szerepe a gyógyászatban és a táplálkozásban [Tin compounds in pharmacy and nutrition]. Acta Pharm. Hung. 2000, 70(2), 53-71.
|
| 68 | Shi, C.; Liu, Z.; Yu, B.; Zhang, Y.; Yang, H.; Han, Y.; Wang, B.; Liu, Z.; Zhang, H. Emergence of nanoplastics in the aquatic environment and possible impacts on aquatic organisms. Sci. Total Environ. 2024, 906, 167404
https://doi.org/10.1016/j.scitotenv.2023.167404 |
| 69 | Shukla, S.; Khanna, S.; Khanna, K. Unveiling the toxicity of micro-nanoplastics: A systematic exploration of understanding environmental and health implications. Toxicol. Rep. 2024, 14, 101844
https://doi.org/10.1016/j.toxrep.2024.101844 |