Enhancing Cellular Immunity Through Epitope Peptides of SARS-CoV-2 in Individuals with Specific HLA Allele
Keywords
Abstract
Background/Aims:
This research explores the effectiveness of a new cytotoxic T-cell epitope peptide specific for HLA-A2402 in enhancing cellular immune responses to SARS-CoV-2 infections. HLA molecules play a key role in presenting antigenic epitopes to T cells, with genetic polymorphisms resulting in varying immune responses among individuals. The study aimed to investigate whether loading this epitope peptide into dendritic cells (DCs) from HLA-A2402(+) individuals could improve immune responses.Methods:
DCs were sensitized with varying doses of the peptide (2–12 µg/mL), with optimal results observed at 8 µg/mL. T-cell responses, proliferation, differentiation, Th cytokine secretion, CTL function, and apoptotic response were compared among the HLA-A2402(+), HLA-A2402(-), PBS control, DC only, and epitope-only groups.Results:
A significant enhancement in DC maturation, antigen presentation, T-cell activation, and proliferation was observed in the HLA-A2402(+) group compared to the HLA-A2402(-) control.Conclusion:
These findings suggest that HLA-A2402-restricted epitope peptides can enhance cellular immunity, offering potential for improving allele-specific SARS-CoV-2 vaccines and other molecular therapies, advancing precision medicine for infectious diseases.Introduction
The immune system consists of several cells and chemicals distributed throughout the body to fend off invaders. Of great importance in this form of defense mechanism is the capacity of T cells to identify foreign bodies. This process includes the T cell receptors interaction with antigenic peptides presented by HLA molecules. The ability of this immune response can be greatly affected by the high degree of polymorphisms found in the HLA gene complex. This genetic diversity is of importance for the ability of the HLA molecules to bind to the antigenic peptides and consequently how the immune system processes antigens to affect T cells and the immune response output strength and quality [1]. For instance, some HLA alleles have higher affinity to some epitopes resulting in more protection against certain pathogens, than those that HLA alleles that do not have such affinities [2, 3]. This variation is one of the key sources of the variation in the immune responses seen across populations and is therefore another area of interest for immunologists and vaccine developers [4].
Three subsets of T cells exist and although all mediate viral clearance cytotoxic T lymphocytes (CTLs) are particularly important in this function [5]. These cells play an important role in maintaining immunity against viral infections such as the COVID-19 [6]. CTLs have played a major role in containing and eradicating SARS-CoV-2 infections [7]. More importantly, the immune response has been shown to vary among individuals and sometimes within the population during the same pandemic [8]. Unfortunately, even with the high coverage vaccination, some appeared to mount a vigorous immunological response while others seemed to have negligible resistance to infection [9, 10]. Although genetic, environmental, and immunological factors might play a part in such variability, HLA polymorphisms that might determine the immunity variance is not fully investigated [11]. Analyzing the genetic factors, such as HLA alleles, required to mount an adequate response to SARS-CoV-2 and subsequently, vaccines, is critical for developing superior therapies and vaccines based on an individual’s genetic profiles [12].
One candidate for this approach is the antigenic determinant peptide VYIGDPAQL, located in the ORF-1ab protein of SARS-CoV-2 [13]. In particular, this peptide has a high binding rate to the HLA-A2402, which is one of the most expressed HLA genes in different populations of the world. Because of such a binding preference, VYIGDPAQL has been put forward for use as an agent that could induce a highly potent CTL response, especially in those expressing HLA-A2402 [14]. Nonetheless, the study finds that this peptide has potential as a CTL activator; however, the objective level of improved immune protection cannot be determined, especially regarding HLA-A2402-positive individuals [15]. This raises a critical question: does the presence of the HLA-A2402 allele in patients correlate with a stronger immune response when exposed to this peptide compared to patients who lack this allele?
In answering this question, this study aims to determine the ability of DCs to promote cellular immunity whenever exposed to the VYIGDPAQL peptide in people with the HLA-A2402 genotype [16]. DCs are central to the activation and regulation of T cells by which they present antigenic peptide fragments to T cells, to instigate an immune response [17]. From studying the aspects of DC maturation, T cell activation, proliferation, and differentiation, the objective of the present study is to investigate if the VYIGDPAQL peptide can specifically augment the immunotherapy in HLA-A2402 positive subjects and thus enhance cellular immunity [18].
These findings have enormous implications given that the ever-emerging diseases and shifts in molecular targets of vaccines require more precise clinical targeting. Should the of VYIGDPAQL peptide strengthen the cellular immunity in patients possessing the HLA-A2402(+) allele, the data obtained can be further used to improve the prospects of a personal vaccine or targeted immunotherapy. It is possible that in the future, vaccines developed to address populations with specific genetic backgrounds through the targeting of HLA could be more effective than current general population vaccines as the immune responses elicited would be unique in each case [19]. Further, the production of peptide-specific T cells could facilitate a rapid and accurate response to immunological threats in susceptible populations within the shortest span [20]. In addition, employing T cells partitioned toward particular alleles of HLA could improve the immunity of such therapies in the long term, as well as protect against newly re-emerging pathogens.
This work also enriches an important but still emerging area of research known as immunogenetics, which is concerned with how genetic differences influence immune responses. As the present research helps specify how HLA polymorphism impacts immune reactions to antigenic peptides, these findings are consistent with the PF campaign to enhance vaccines and immunotherapies [21, 22]. Vaccines and therapies could be developed to target the different HLA types in various human populations would not only enhance the general effectiveness of any treatment but also level out immune discrimination across populations [23]. Hence, our strategy to examine how the VYIGDPAQL peptide affects immune responses in HLA-A2402(+) individuals is crucial to improving medical therapies.
Finally, this work aims to offer the basis for the application of immunogenic discoveries to prevent and control infectious diseases, with a focus on SARS-CoV-2 and other emerging viruses. This could help define future novel vaccines or immunotherapy, and make it possible to design treatments that are personalized to a participants’ genomics type, thus potentially improving health systems results or survival rates globally. By understanding how HLA polymorphism influences immune response, we move closer to a future where personalized genomic treatments can effectively combat disease-causing pathogens and control infectious agents.
Materials and Methods
Experimental Materials
Fig. 1: Instrumentation and Reagents.
Experimental Methodology
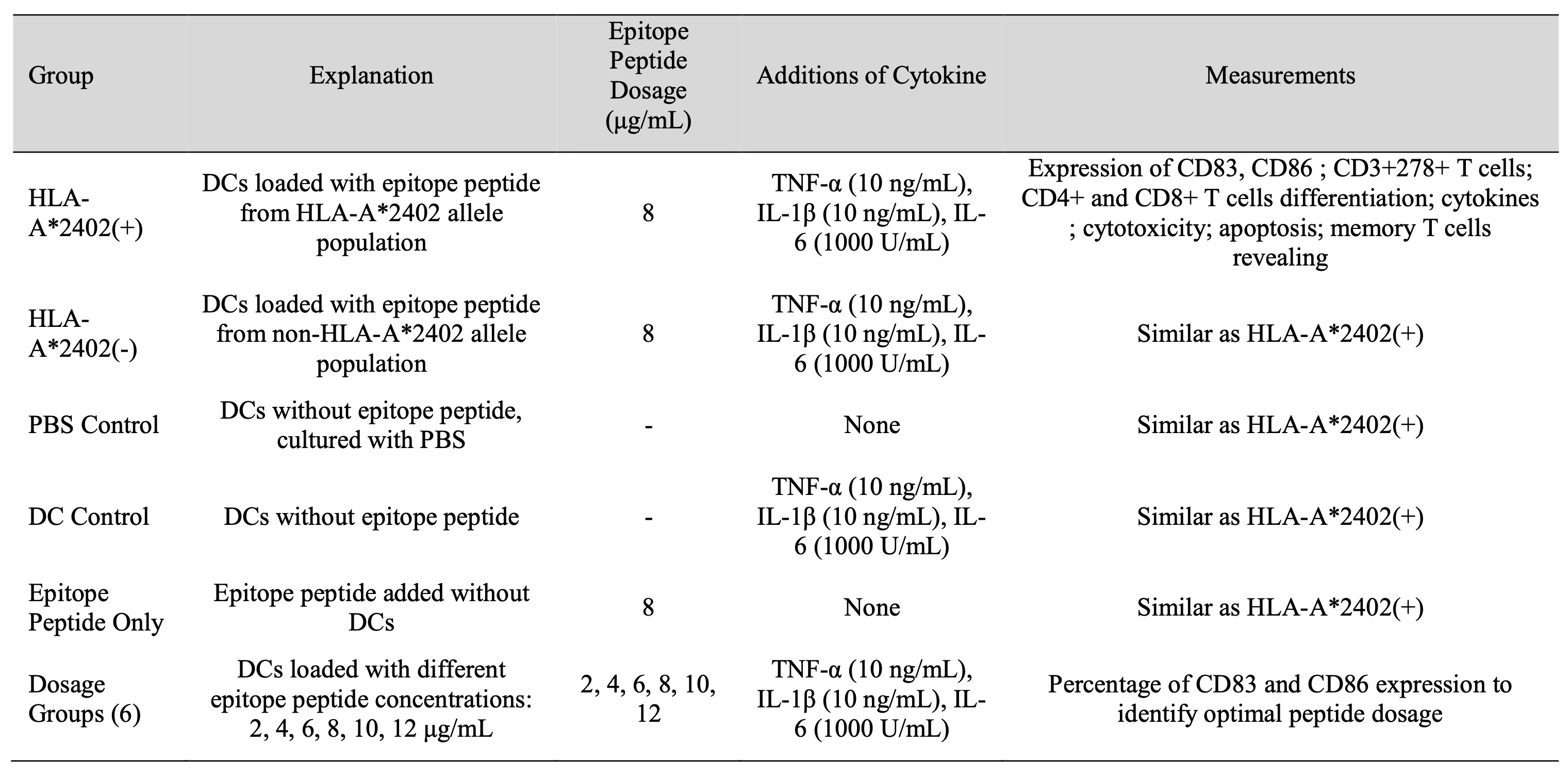
Table 1: Trial Groups and Epitope Peptide Treatments

Table 2: Trial Assays and Analytical Approaches. Table 2. outlines the experimental procedures and analytical techniques used to assess the effects of antigenic determinant peptides on immune response modulation. The assays include dendritic cell (DC) activation, antigen presentation, T-cell activation and differentiation, cytokine secretion, cytotoxicity assessment, apoptosis evaluation, and memory T-cell formation. Flow cytometry, MTT assays, LDH assays, and Luminex cytokine profiling were used to quantify these immune parameters.
Results
Dose-dependent maturation of DCs loaded with epitope Peptides
The results showed that the density of CD83 and CD86 on DCs increased in a dose-dependent manner, the highest density of these receptors observed at 8 µg/ml of the epitope peptide. This concentration was associated with the highest level of natural DC maturity as evaluated by flow cytometry compared to other concentrations (2, 4, 6, 10, and 12 µg/mL). The concentration of 8 µg/mL was used in all the following experiments to enhance the initiation of cellular immune response. These results underline the necessity to increase the doses of the concentration to increase the efficiency of DC antigen presentation, which can significantly improve the productivity of immune response (see Table 3).
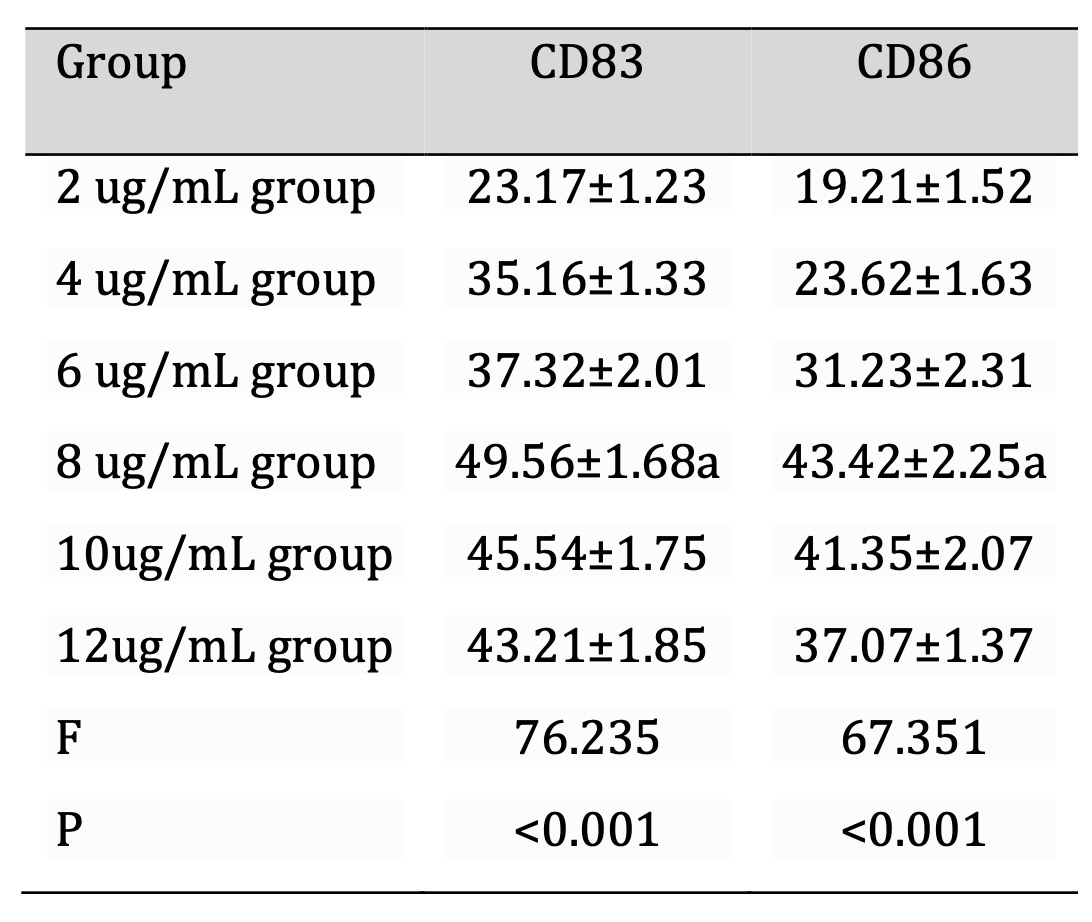
Table 3: Percentage of DCs expressing CD83 and CD86 in each group (%, x±s, n=30). Note: DCs were cultured in AIM-V medium and treated with epitope peptides at different concentrations for 8 days. Flow cytometry analysis was performed using FITC-CD83 and FITC-CD86 antibodies to assess DC maturation. Data were acquired using a BD Accuri C6 cytometer, and gating was applied to exclude debris. Statistical analysis was conducted using one-way ANOVA followed by Tukey's post hoc test. Values represent mean ± standard deviation (SD), with significance set at P < 0.05.
Increased DC Antigen Presentation in HLA-A * 2402 (+) population
We observed that HLA-A2402 significantly increases the levels of CD83 and CD86 in DCs. HLA-A2402 (+) group exhibited much higher levels of both CD83+ and CD86+ compared to HLA-A2402 negative (-) and PBS control. These observations were obtained at a significant level (n = 15, P<0.05) showing a significant relationship between the presence of HLA-A2402 and high activation markers on DCs. This implies that HLA-A2402 (+) may play a role in the modulation of DC immune functions possibly by strengthening the overall immune response. Further elaboration of the input conditions for this observation is presented in Table 4.
Table 4: Percentage of DCs expressing CD83 and CD86 in each group (%,`x±s, n=30). Note: DCs were loaded with 8 µg/mL of the epitope peptide and co-cultured with PBMCs from HLA-A2402(+) and HLA-A2402(-) groups for 8 days. Flow cytometry analysis was performed using FITC-CD83 and FITC-CD86 antibodies to assess DC maturation. Data acquisition was carried out using a BD Accuri C6 cytometer, with gating applied to exclude debris. Statistical comparisons were conducted using one-way ANOVA followed by Tukey's post hoc test. Values represent mean ± standard deviation (SD), with significance set at P < 0.05.
T Cell Activation (CD3+278+)
The immunogenicity of epitope peptide in association with the HLA-A2402 molecule was determined by T-cell activation across different groups. The HLA-A2402 (+) group revealed a more marked T cell activation than the HLA-A2402 (-) group, as well as other control groups of PBS, DC, and epitopes. In particular, the HLA-A2402 (+) group underwent a higher index of the immune activation marker CD3+CD278+ T cells, which suggested the importance of HLA-A2402 in the presentation of the peptide and the subsequent initiation of effective immune response. Comparison of all groups also showed that the percentage of CD3+CD278+ T cells was significantly lower (P < 0.05) in the participants group with HLA-A2402 (+) than in all the other groups. The experimental results of flow cytometry which support the T cell activation pathway (Fig. 2A and 2B). Based on these observations, it is conceivable that the Ag-3RP3 could be employed for designing antigen-specific immunotherapeutic strategies for HLA-A2402 (+) individuals. (Table 5, Fig. 2)
Note: T-cell activation was assessed using flow cytometry on day 10 after co-culture with dendritic cells. CD3⁺278⁺ (ICOS) expression was measured using PE-CD3 and FITC-CD278 antibodies. Data acquisition was performed using a BD Accuri C6 cytometer, with gating applied to exclude doublets and debris. Group comparisons were conducted using one-way ANOVA followed by Tukey’s post hoc test. Values represent mean ± standard deviation (SD), with significance set at P < 0.05.
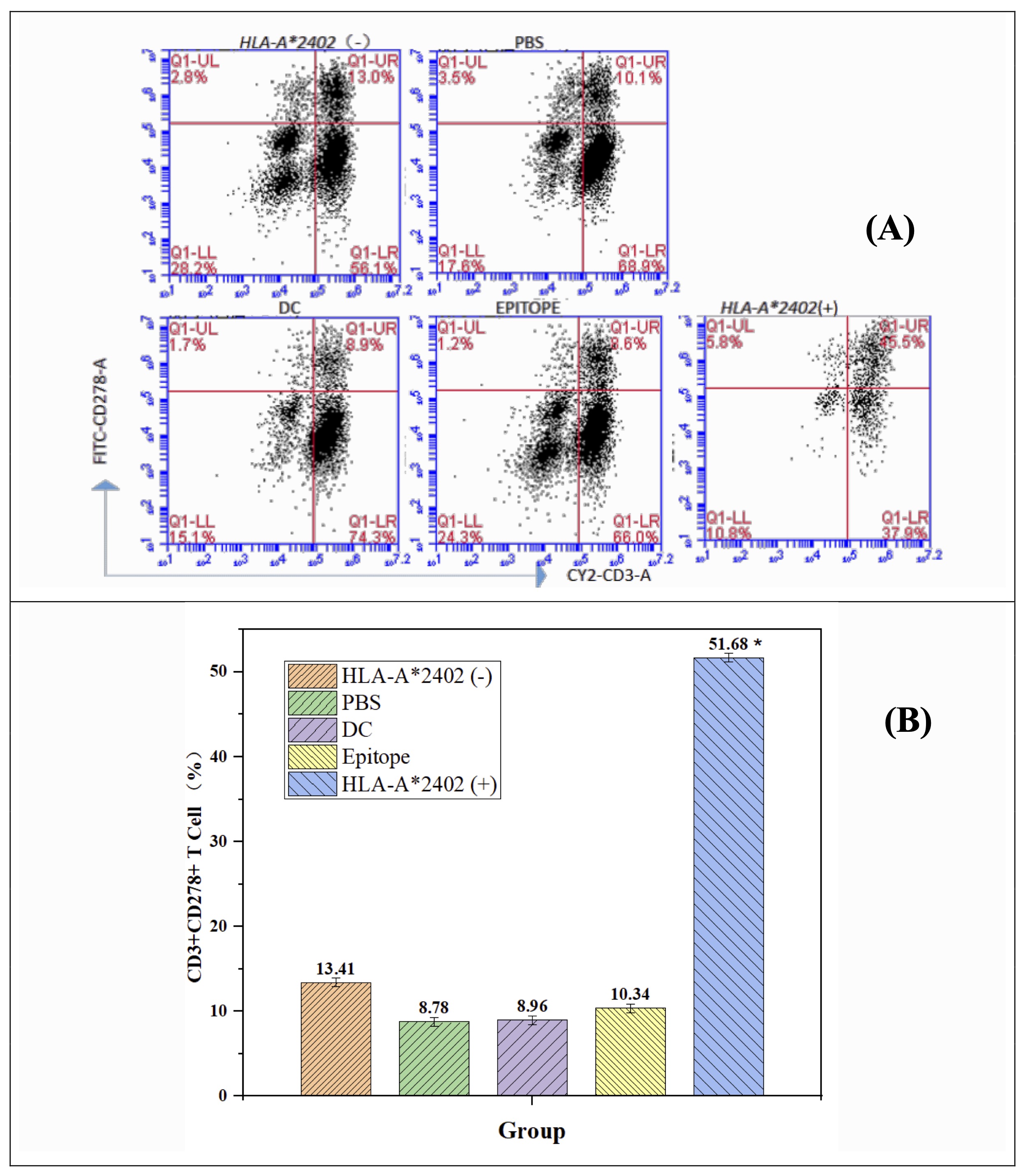
Fig. 2: DC-loaded epitope peptides from the HLA-A*2402 allele population promote auto T cell activation. Note: n = 15, F = 73.162, p < 0.001, * representing a major difference when compared with the HLA-A2402(-), PBS, DC, and Epitope groups.
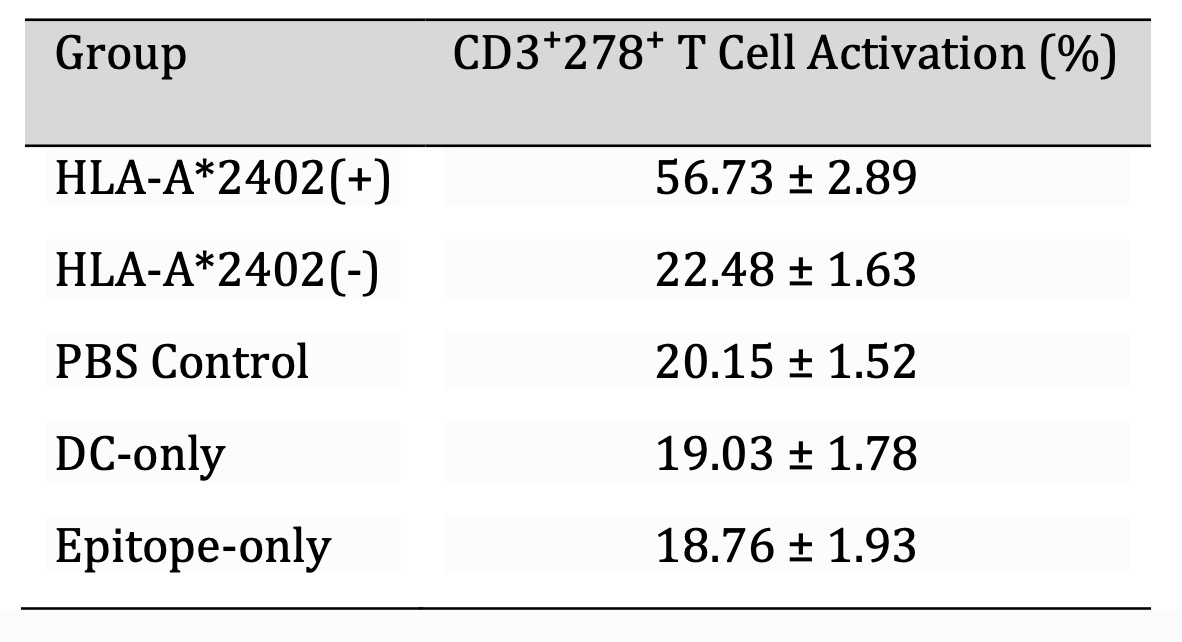
Table 5: Percentage of CD3⁺278⁺ T Cell Activation in each group (%,`x±s, n=30). Note: T-cell activation was assessed using flow cytometry on day 10 after co-culture with dendritic cells. CD3⁺278⁺ (ICOS) expression was measured using PE-CD3 and FITC-CD278 antibodies. Data acquisition was performed using a BD Accuri C6 cytometer, with gating applied to exclude doublets and debris. Group comparisons were conducted using one-way ANOVA followed by Tukey's post hoc test. Values represent mean ± standard deviation (SD), with significance set at P < 0.05.
DC-Loaded Epitope Peptide from the HLA-A2402 Allele Population Enhances Proliferation of Autologous T Cells
In this study, the proliferative function of T cells of HLA-A2402 (-), PBS, DC, and epitope peptides has been compared concerning different experimental groups. There was a significant difference (p<0.05) in the present study, stressing the immunologic role of HLA-A2402. These results indicate that HLA-A2402 might improve immune activities and provide a better understanding of T lymphocytes in immune-regulatory processes. Additional analysis gives a more detailed look at these discrepancies, which confirm that HLA-A2402 has a central role in regulating immune response (Fig. 3).
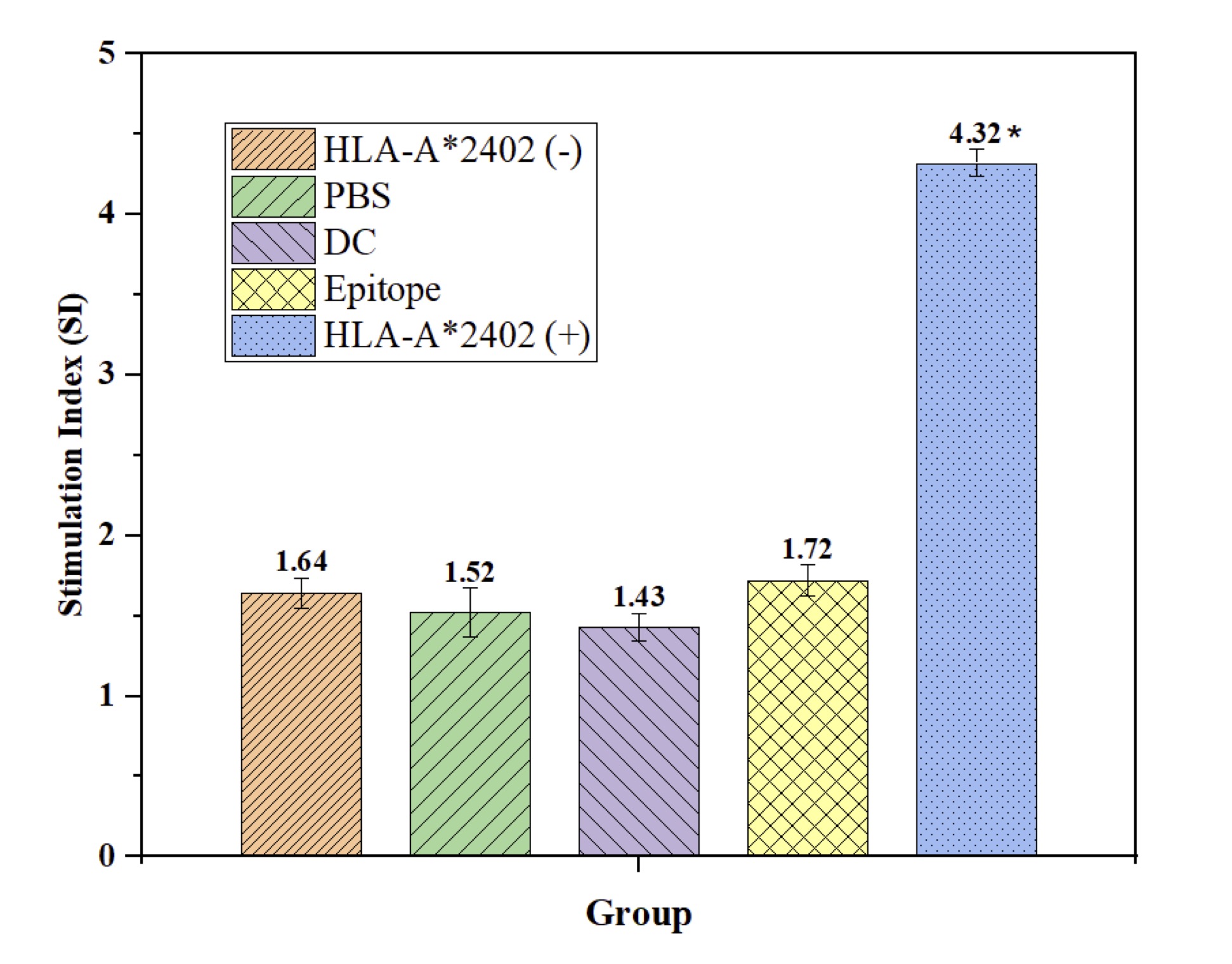
Fig. 3: DC-Loaded Epitope Peptides from the HLA-A2402 Allele Population Promote Self T Cell Proliferation. Note: n = 15, F = 73.162, p < 0.001, * compared to HLA-A2402(-), PBS, DC, and Epitope-only groups.
DC-Loaded Epitope Peptide from the HLA-A*2402 Drives Expansion of CD8+ Cytotoxic T Cells
In this study, we evaluated the effects of HLA-A2402 on the percentage of CD8+ and CD4+ T cells within different experimental setups. It was shown that the HLA-A2402(+) group had a higher count of CD8+ T cells compared with the HLA-A2402(-) group, PBS, DC, and epitope peptide groups with P < 0.05, as shown in Fig. 4. This implies that specifically, the HLA-A2402 serves a very important role in boosting the CD8+ T cell immunological response. On the other hand, the proportion of CD4+ T cells showed significant differences between the HLA-A2402(+) group and other groups (HLA-A2402(-), PBS, DC, or epitope peptide) with no effect on the proportion of CD4+ T cells. These findings show that HLA-A2402 is highly selective in altering immune response, favoring CD8+ T cells while having little effect on CD4+ T cells. Hence, HLA-A2402 would seem to play an essential role in controlling the activation and proliferative activity of cytotoxic T lymphocytes in immunological terms that would reassert the molecule’s importance in immune response modulation (Fig. 4).
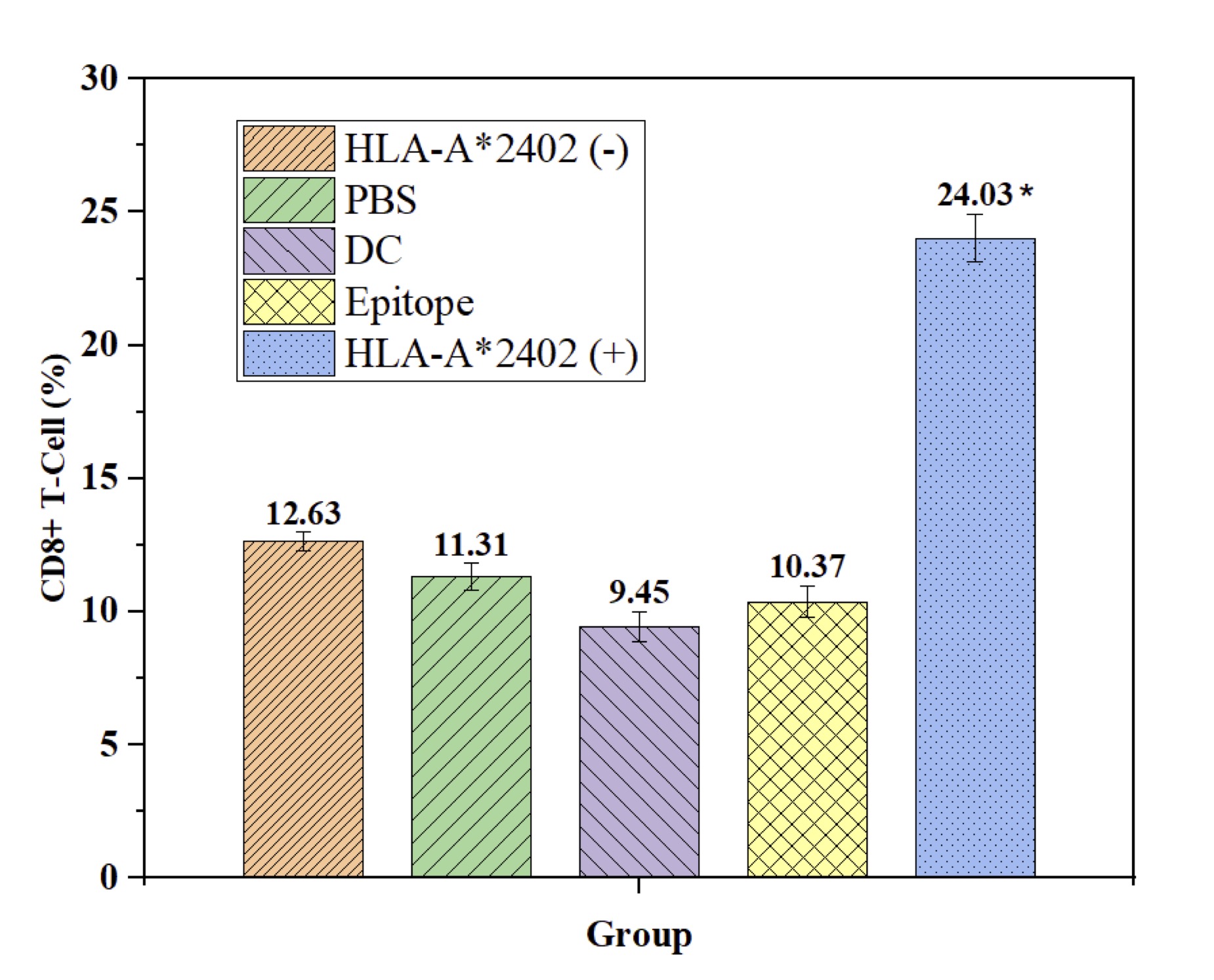
Fig. 4: DC-Loaded Epitope Peptides from the HLA-A2402 Allele Population Promote Differentiation of Self T Cells into CD8+ T Cells. Note: n = 15, F = 24.346, p < 0.001, * compared with HLA-A2402(-), PBS, DC, and Epitope groups.
Investigation of Th Effects Induced by DC-Loaded Epitope Peptides in the HLA-A2402 Allele Population
The present work aims to investigate the T-helper (Th) response to human dendritic cell (DC)-loaded antigenic peptides that may be recognized by HLA-A2402. The research compared five different groups: HLA-A2402(+) subjects and HLA-A2402(-) subjects, healthy controls, and the effects of PBS-treated samples, DC-loaded samples, and epitopic peptides. To test differences in immune response among these groups, cytokines IL-2, IL-5, IL-10, IL-12p70, GM-CSF, IFN-γ, and TNF-αwere measured as shown in Table 6. In addition, there were also no significant differences in the level of cytokines released in. From these results, it has been inferred that the HLA-A2402 allele does not have a large and influential part in the management of the Th immune response evoked from the peptide bound to the DC. More work has to be done to analyze the pathways and find out the possibilities that are linked to HLA-A2402 as an immunotherapeutic target in limited ethnic groups (Table 6).
Table 6: Comparison of Cytokine Expression in HLA-A2402(+) and HLA-A2402(-) Groups. Note: Cytokine levels (µg/mL) were measured using a multiplex Luminex assay across five experimental groups: HLA-A2402(+), HLA-A2402(-), PBS control, DC-loaded samples, and epitope peptide-only samples. Statistical analysis was performed using one-way ANOVA followed by Tukey’s post hoc test, with no significant differences observed between HLA-A2402(+) and HLA-A2402(-) groups (p > 0.05).
Improvement of Cytotoxic T Lymphocyte Activity via Dendritic Cell-Loaded Epitope Peptides in HLA-A2402(+) Populations
This study focuses on whether the expected antigenic determinant peptides provided onto dendritic cells can increase cytotoxic T lymphocyte response among the HLA-A2402 carriers. According to the real experimental data, the HLA-A2402(+) group had a higher OD value of cytotoxicity as compared to the HLA-A2402(-) group, PBS, DC-only, and epitope peptide-only groups (p< 0.05). Similarly, the cytotoxic outcome was significantly higher in the HLA-A2402(+) group when the effector/target (E/T) ratio was 100:1 (p< 0.05). However, no significant differences were found when the E/T ratio was 25:1. These results underscore the possibility of employing DC-loan antigenic peptides that would stimulate CTL cytotoxic activity in HLA-A2402(+) subjects, making possible the treatment of immunological diseases that are HLA-restricted (Fig. 5).
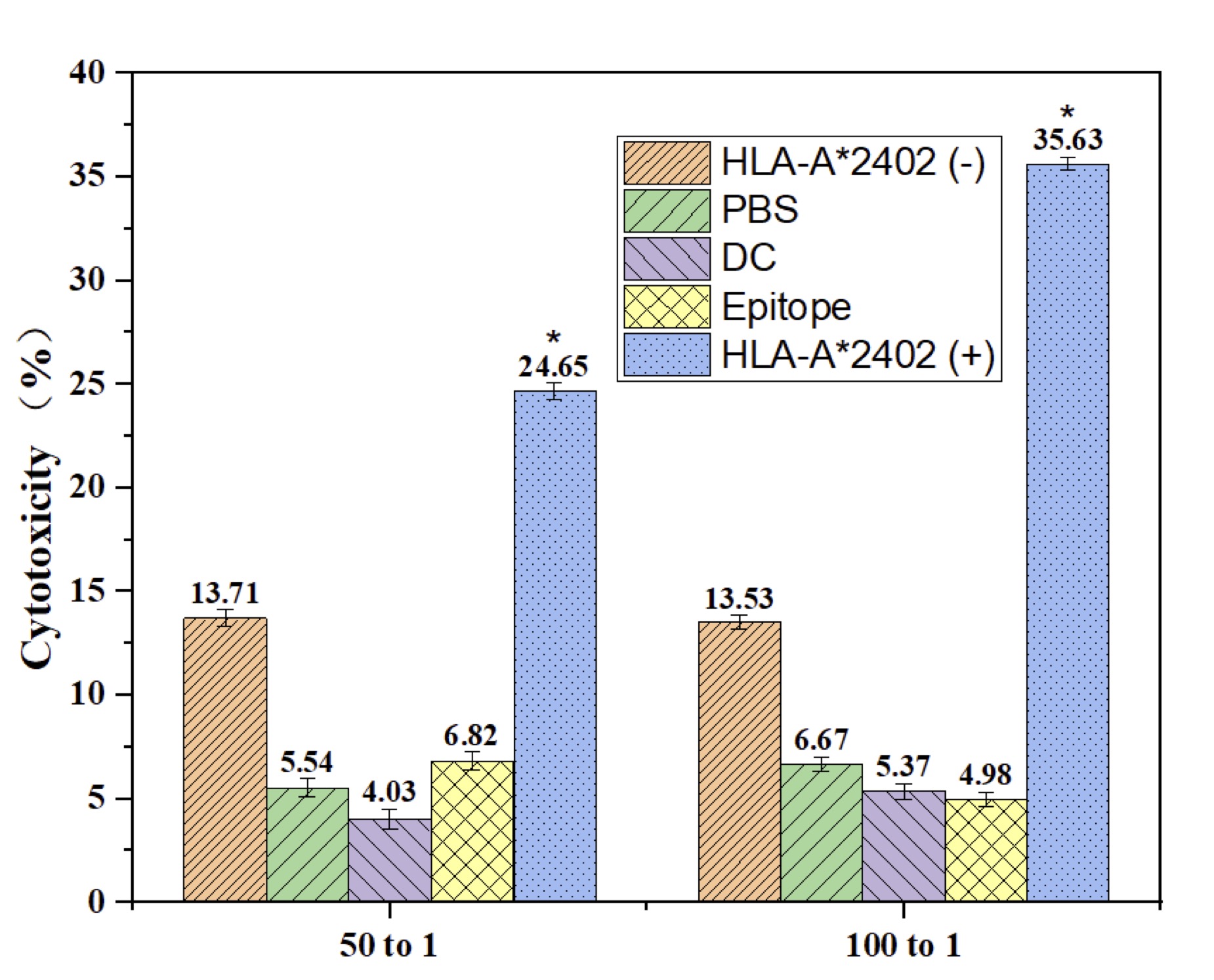
Fig. 5: Cytotoxic Effects of DC-Loaded Epitope Peptides from the HLA-A2402 Allele. Note: n = 30, F = 17.233, p > 0.05. * DC-loaded epitope peptides showed a trend toward reduced apoptosis compared to the HLA-A2402(-), PBS, DC, and Epitope-only groups; however, the differences were not statistically significant.
Epitope Peptide-Loaded Dendritic Cells Induce Apoptosis in HLA-A*2402(+) Populations with Important Selectivity.
Here, we review the influence of antigenic determinant peptide-loaded dendritic cells on apoptosis in patients with HLA-A2402. The results indicated that a smaller proportion of apoptotic cells in the HLA-A2402 (+) group compared with HLA-A2402 (-), PBS-treated, DC-alone, and epitope peptide-alone groups (p<0.05). This demonstrates a selective apoptotic effect, which is elicited by the presence of the HLA-A2402 allele, signifying its role in peptide-mediated immune stimulation. The study also provides insights into how understanding the specific HLA genotype may increase the accuracy and efficacy of cancer immunotherapy and serves as the groundwork for even further developments of new targeted therapy (Fig. 6).
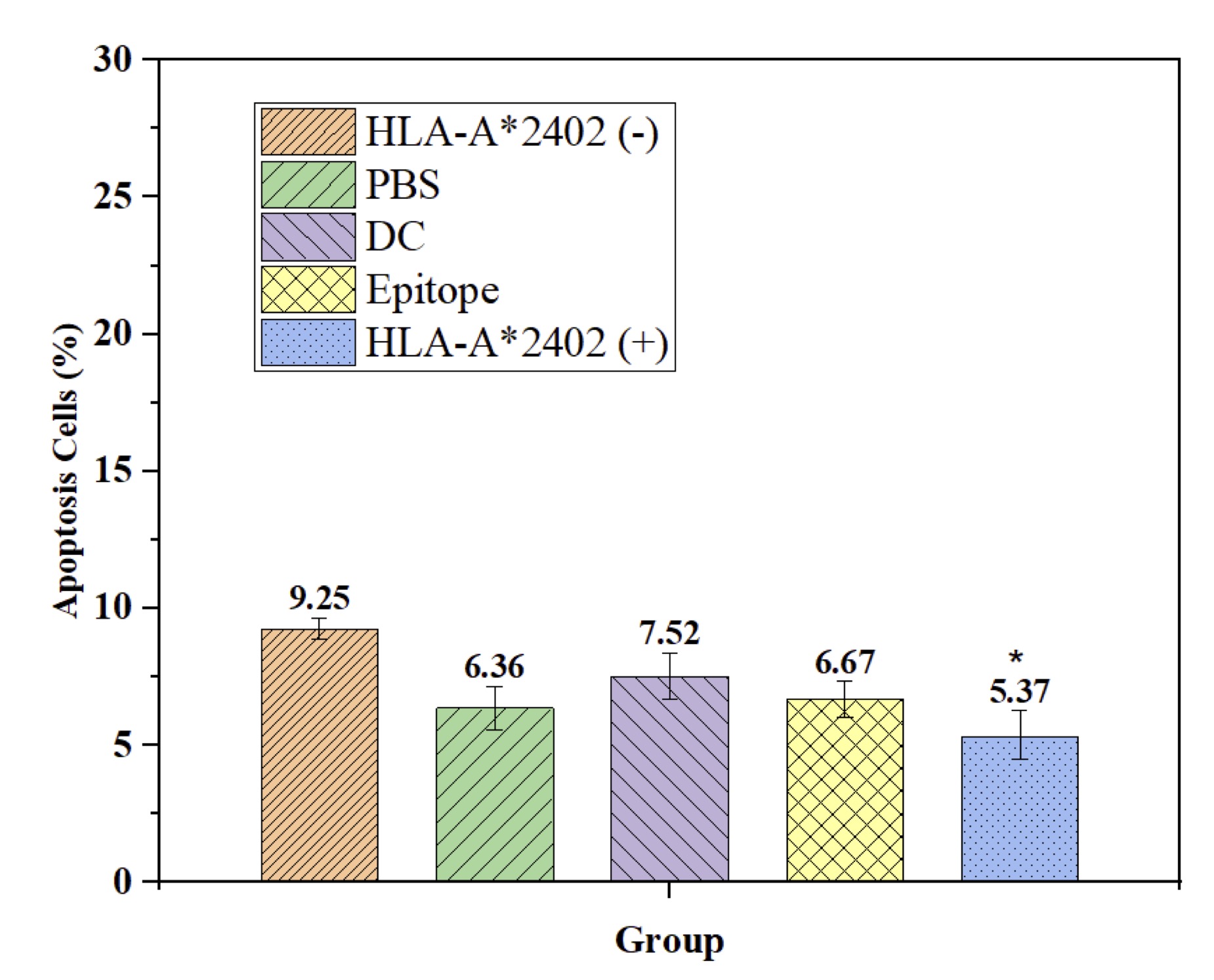
Fig. 6: Reduced Apoptosis Induced by DC-Loaded Epitope Peptides in the HLA-A*2402 Allele Population. Note: n = 15, F = 7.213, p < 0.001, * compared with HLA-A2402(-), PBS, DC, and Epitope groups.
DC-Loaded Epitope Peptides from HLA-A*2402+ Populations Enhance CD8+ Memory T Cell Differentiation
This study focuses on the effects of peptides incorporating determinant antigens from HLA-A2402(+) populations on the formation of CD8 + memory phenotype T cells. Compared in HLA-A2402(-), PBS group, DC group, and epitope peptide group, CD8+ Memory T cell of the HLA-A2402(+) group is significantly higher (p<0.05). This suggests that there is a strong requirement for the processing of HLA-A2402-specific antigenic peptides by DCs in the course of CD8+ memory T cell maturation. Surprisingly, no statistically significant difference was observed in the percentage of the CD4+ memory T cells (p>0.05); the findings of this study underlined that the chosen antigenic peptide influences the CD8+ T cells. These results offer great promise to guide future strategies in vaccine and immunotherapy design by directly targeting the CD8+ T cell population to augment immunity against diseases (Fig. 7A and 7B).
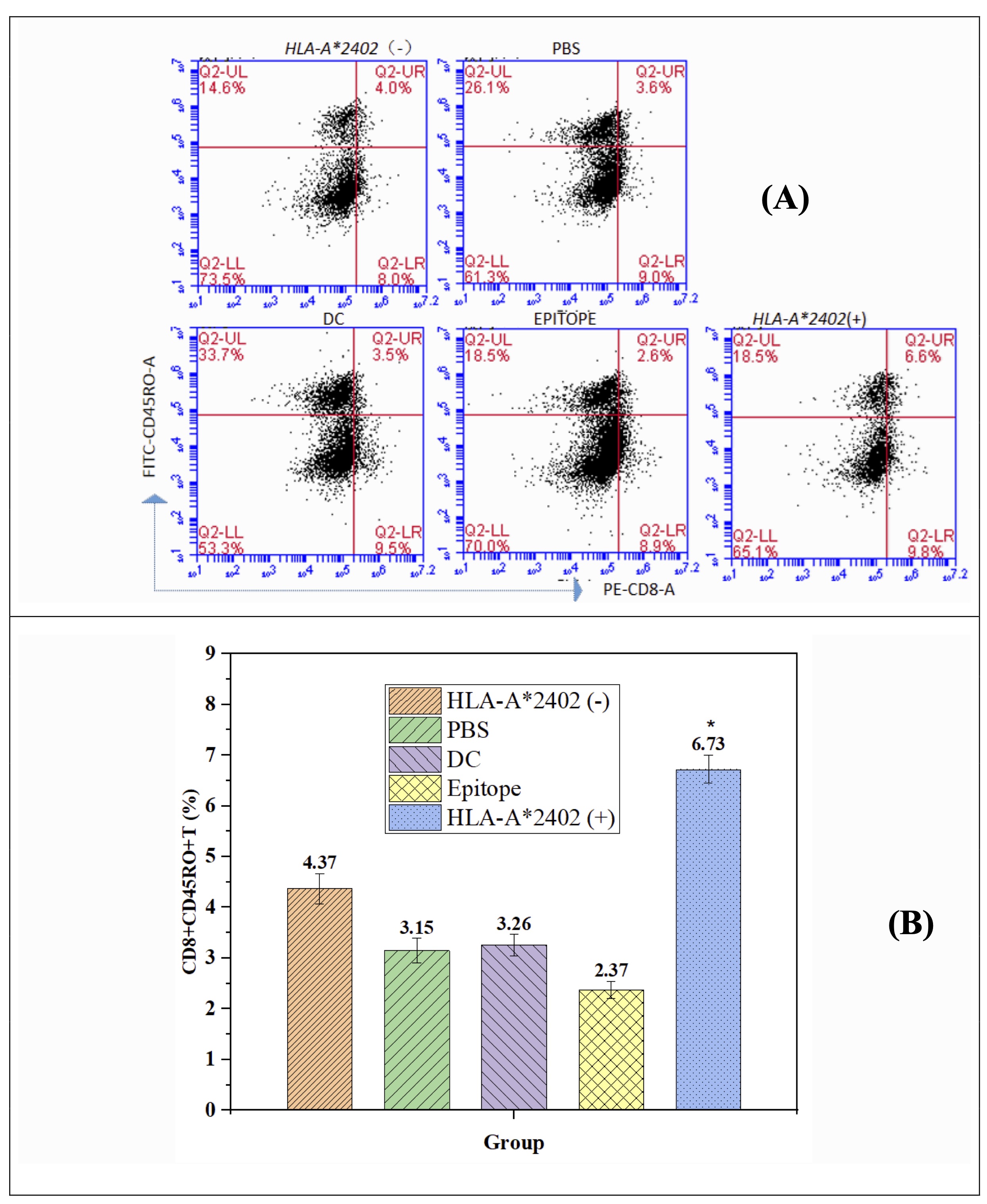
Fig. 7: DC-Loaded Epitope Peptides from the HLA-A2402 Allele Population Promote CD8+ Memory T-Cell Formation. Note: n = 15, F = 4.673, p < 0.001. * Meaningfully different from HLA-A2402(-), PBS, DC, and Epitope groups.
Immunological Improvements via HLA-A*2402-Restricted Epitope Peptide: Perceptions from DC-Mediated T Cell Activation and Participant Demographics
This work addresses the immunological properties of the antigenic determinant peptide based on the HLA-A*2402 and its ability to induce differentiation of auto-aggressive CD8+T cells. The results offer a general understanding of how this peptide increases cell permeability via the DC-dependent pathway, increasing T lymphocyte activation, proliferation, and cytotoxicity. The study also focuses on epitope-based vaccines as the future of precise immunotherapy, which confirms the value of targeting the right antigens in the immune response.
The study compares several experimental groups: HLA-A2402 (+) loaded with a peptide derived from DC, HLA-A2402 (-) received PBS instead, epitope, only DC, and epitope only. A marked enhancement in the percentage of expression of CD83/CD86 was observed in the HLA A-2402(+) group at 48.73% and 45.68%, respectively, indicating enhanced maturation of DC. However, the HLA-A2402 (-) group had a lower rate of maturation (21.63% and 24.27%). Furthermore, the HLA-A2402 (+) group presented higher T cell activation characterized by a higher stimulation index (SI=5.32±0.34) than the other groups. This group also expressed high cytokine levels representative of the active immune response, while the other groups expressed low cytokine production, showing that this peptide is highly potent in activating the immune response (see Table 7).
The cytotoxicity of the HLA-A2402 (+) group with 78.42 ± 3.14 % of the cells was significantly higher in comparison with other groups: HLA-A2402 (-) with 34.18 ± 2.57 % as well as PBS direct group with 22.53 ± 1.76 %. In the same vein, the HLA-A2402 (+) people also had significantly significantly low apoptotic levels (8.23 ± 1.02 %), an indication that immune cells in this group were more efficient and durative. The outcomes of these studies reveal confirmed the high level of immunity that is linked to the HLA-A2402 (+) allele, which should be favorable for the treatment of this disease (Fig. 7).
Specifically, in terms of demography, there was no difference between the HLA-A2402 (+) and the HLA-A2402 (-) participants. The participants’ ages varied from 20 to 40 years; the mean ages were 31.5 ± 4.8 years in the HLA-A2402 (+) group and 32.2 ± 5.1 years in those of the HLA-A2402 (-) group, decreasing any potential age-related prejudice. This variable was equally split, but males had a slight majority in both groups. It was an ethical study, and all participants’ safety and free will were obtained from the institutional review board.
Population and sample characteristics and ethical practices show that the study has the reliability of data collected and analyzed. The combined data provides a better understanding of HLA-A2402-restricted antigenic peptides to give the basic knowledge on how to further develop vaccines that improve the immune system response in autoimmune diseases and Cancer therapies (see Table 8).
This study brings important emphasis to the fact that cellular insusceptibility involves the HLA-A*2402-restricted antigenic determinant peptide, suggesting that this occurs through the pathway activated by DC. The observed increase is most probably a result of improved activation, proliferation, differentiation, cytotoxicity, and memory formation of T cells. Overall, these results provide a strong validation of the efficacy of epitope-based vaccines in precision immunotherapy since the epitopes can selectively induce certain types of reactions.
Table 7: Comparison of Experimental and Control Groups. Note: DC maturation was assessed by flow cytometry using FITC-CD83 and FITC-CD86 antibodies on a BD Accuri C6 cytometer. T-cell proliferation was measured via MTT assay at 570 nm after a 4-hour incubation. Cytokine levels (IFN-γ, IL-2) were quantified using a multiplex Luminex assay. Cytotoxic effects were determined by LDH release assay at 492 nm after 24-hour incubation. Apoptosis was analyzed via Annexin V-PE and PI staining, with flow cytometry used to distinguish apoptotic cells. Statistical comparisons were conducted using one-way ANOVA followed by Tukey’s post hoc test, with significance set at P < 0.05.
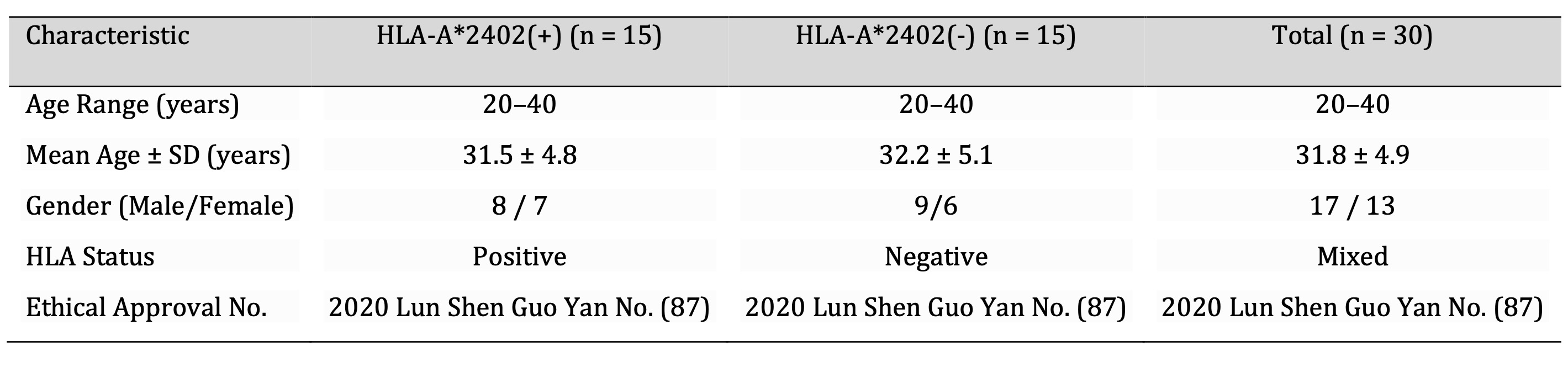
Table 8: Participant Demographics. Note: An equal number of participants (n=15 per group) were recruited to maintain genetic variability and reduce selection bias. Age and gender distribution were balanced to ensure comparability between HLA-A2402(+) and HLA-A2402(-) groups. Ethical approval was obtained (Approval No. 2020 Lun Shen Guo Yan No. 87), and all participants provided informed consent in accordance with institutional and international ethical guidelines.
Discussion
Most SARS-CoV-2 vaccines are developed based on the receptor binding domain (RBD) of the viral spike protein [24]. However, it was seen that there was a decrease in titers of antibodies after approximately three months of post-vaccination, these vaccines are claimed to provide decreased long-term immunity [10], [24]. In contrast, SARS-CoV-2-specific CD8+ T cells have shown the possibility to prolong the immune memory and the mild symptoms of diseases in animals; this suggests that T cell immunity could provide extra protection [25, 26]. Such considerations illustrate the importance of extending the characteristics of vaccines to include T-cell immune responses. In this study, we investigate the immunological features inherent to the SARS-CoV-2 antigenic peptide restricted to HLA-A2402. It has good binding to this HLA-A2402 molecule, which is produced in some population groups. Experimental data in vitro indicate that the peptide has a positive impact on the dendritic cell (DC)-mediated processing of antigens with subsequent activation of cellular immunity, the generation of effector, and memory CD8 T cells. In light of these findings, the epitope-based vaccines specific to the HLA-A2402 allele are a helpful guide for the generation of cellular immunity against SARS-CoV-2. SARS-CoV-2 vaccines cause different immunologic reactions against the virus. The consideration of antibody-mediated immunity as constitutive of the total immunity response is important as they contribute 40% in countering the virus and offer an immediate solution by virtually immobilizing the virus. 35% of cellular immunity is played by CD8+ T cells through the killing of infected cells and long-term immune memory. The highest, 15% of CD4+ T cell support, improves the immune response by helping both antibodies and T-cells. They noted that 10% of this defense system consists of other immune mechanisms that offer support enhancement. These responses sum up the confusion regarding immunization induced by vaccines, thus suggesting that different mechanisms should be addressed to achieve long-lasting immunity (Fig. 8A, B).
Fig. 8: Immune Responses and Advantages of SARS-CoV-2 Vaccines; (a): Immune Responses to SARS-CoV-2 Vaccines; (b): Vaccine Advantages Against SARS-CoV-2. (This figure was generated by the author based on the study's findings).
Implications for Vaccine Development
Epitope-based vaccines seem to have certain advantages, the most attractive being the capacity of the vaccines to stimulate a precise and highly functional CD8+ T cell response. Unlike antibody-referenced immunity, these vaccines can overcome the problems posed by mutations in SARS-CoV-2 due to conserved T-cell antigens. The current study focuses on the subject of loading dendritic cells (DCs) with HLA-A*2402-restricted antigenic peptides and the effects on T-cell activation, proliferation, and differentiation. This finding concurs with results advanced by previous studies suggesting the viability of DC-based immunotherapy when used to address different infectious or malignant diseases. In addition, the generation of memory CD8+ T cells will confirm that this approach will offer advantageous and durable protection, which is crucial for the safety of a vaccine. Altogether, high T cell immunogenicity and long-lasting immunity imply that epitope vaccines could provide sufficient protection against new viral variants and maintain protective immunity [27]. Epitope-based SARS-CoV-2 vaccines present several essential features that should be prioritized. Cell-mediated immunity, accounting for 35%, contributes to long-term immunity by generating memory cells. Explaining mutations, which account for 30 %, ensures these vaccines are effective against new strains to do the usage of T cell antigen. Memory T cell generation, which is an advantage that contributes 25% as shown in Fig. 8B, provides long-term immunity and reduces disease severity. Other benefits of taking an abridged 10% are increased safety and specific measures. These benefits have made epitope vaccines the most useful instrument for further immunization campaigns, as they are not only effective instantly but also easily adaptable to the evolving landscape of viral mutations (see Fig. 8B).
Optimizing Vaccine Delivery and Immune Responses
Analyses have shown that the success of an epitope-based vaccine depend not only on choosing a good, high-affinity peptide but also on how that peptide is presented. DC is defined as an essential platform for the cross-presentation of HLA class I molecules to activate the CD8+ T cells. Also, regarding mucosal immunity, alveolar epithelial cells facilitate the vaccine’s effectiveness [28]. Further enhancement of the ability to present antigenic peptides to CD4+ T cells by the combination of adjuvants such as TLR antagonists and exosomes was proposed [29]. This approach may also expedite the development of immunity, a critical factor in combating and containing the pandemic. In addition, there should be an emphasis on promoting the survival of memory T cells. CD4+ T-cells have been found in patients who recovered from SARS-CoV-2 infection, the effector memory T cells, associated with intense immune response [25, 30]. Another very active area of research is tissue-resident memory T cells, which provide site-specific immunity. This is an important area of research as the interactions between the memory cell subsets and how they contribute to the overall goal of long-term immunity need to be fully comprehensible to enhance the development of vaccines.
Study Limitations and Future Directions
However, some problems must be solved before the results of the investigation are confirmed. This was the first experiment that was in vitro , meaning that it did not reflect real-life results of what happens within the body (in vivo ). For the confirmation of the effectiveness and safety of the proposed methods of vaccination in large-scale situations, animal models and clinical trials are the logical next step. Further, the present study is restricted to the HLA-A*2402 allele only, which reduces the generalizability of the study population. However, the variability of HLA alleles has a great impact on the immune responses to infections; therefore, developing new studies to identify relevant antigenic determinant peptides to other HLA alternatives. It would increase the capacity of corrective immunotherapy even more.
Even implementing these findings into actual practice comes with several great challenges. One major concern is how to scale up dendritic cell (DC) vaccine production; in other words, there is a need to find effective and standardized methods for loading dendritic cells with the appropriate peptides. Further improvements in the fabrication methods, as well as cost-cutting measures, will be required to scale up these therapies. Last but not least, the study presses the point about the need to conduct long-term research in the handling and improvement of vaccines. It has been suggested that the conjugation of antigen peptides with adjuvants, including TLR agonists, may amplify immune responses. Additionally, the combination of epitope vaccines with the most effective methods of delivery today, such as mRNA or nanoparticles, could enhance their immunogenicity.
Conclusion
The current study substantiates our previous hypothesis that the DC-based load of the HLA-A*2402-restricted SARS-CoV-2 antigenic determinant peptide improves the critical immune response profiles such as CD8+ T cells, memory T cell generation, and reduced apoptosis level, thus increasing cell permissiveness to SARS-CoV-2 in vitro . These data support the ability of epitope-based vaccines to circumvent the issue observed in current SARS-CoV-2 vaccines where the population with specific HLA alleles has demonstrated lower efficacy. The epitope-based approach is, therefore, a specific approach that concerns the immune responsiveness in such populations.
However, several issues are worth capitalizing on but there are a few things that should also be taken into consideration. Some of the drawbacks of this study are the selection of the second in vitro environment, restriction of the subject demographics, and difficulty in generalizing the findings to practice scenarios, which require further research. Further investigations of the subject should incorporate more HLA alleles, in vivo models should be involved, and other treatment regimens should be co-administered with adjuvants.
Finally, the epitope-based vaccine strategy has valuable potential to enhance immunity not only against SARS-CoV-2 but also against many other novel microorganisms. This should open up new opportunities for optimizing vaccines for their ability to work well as a treatment and protection against different diseases and for various population groups/variants, thus improving results of combating the infections.
Acknowledgements
Author Contributions
X.H.: conceptualization, methodology, investigation, writing the original draft.
Y.W.: methodology & investigation. F.Y.: writing, review & editing, project administration, funding acquisition, supervision. B.D.: methodology & investigation.
Z.Y.: conceptualization, writing, review & editing, supervision.
Funding Sources
Anhui Provincial Department of Education Natural Science Research Project Major Project (2023AH040263), Anhui Provincial Department of Education Natural Science Research Project Key Project (KJ2019A0415, KJ2021A0829), Wuhu Science and Technology Program (No. 2020ms3-9), Wannan Medical College Scientific Research Project (No. wyqnyx202004 and WK2021Z07).
Statement of Ethics
Institutional Review Board Statement: NA
Informed Consent Statement: Informed consent was obtained from all subjects involved in the study.
Disclosure Statement
The authors declare no conflicts of interest. No AI tools have been used to create this work.
References
| 1 | Khan T, Rahman M, Ahmed I, Al Ali F, Jithesh PV, Marr N. Human leukocyte antigen class II gene diversity tunes antibody repertoires to common pathogens. Front Immunol. 2022 Aug 8;13:856497. doi: 10.3389/fimmu.2022.856497. PMID: 36003377; PMCID: PMC9393332.
https://doi.org/10.3389/fimmu.2022.856497 |
| 2 | Barquera R, Collen E, Di D, Buhler S, Teixeira J, Llamas B, Nunes JM, Sanchez-Mazas A. Binding affinities of 438 HLA proteins to complete proteomes of seven pandemic viruses and distributions of strongest and weakest HLA peptide binders in populations worldwide. HLA. 2020 Sep;96(3):277-298. doi: 10.1111/tan.13956. Epub 2020 Jun 11. PMID: 32475052; PMCID: PMC7300650.
https://doi.org/10.1111/tan.13956 |
| 3 | Mosaad YM. Clinical Role of Human Leukocyte Antigen in Health and Disease. Scand J Immunol. 2015 Oct;82(4):283-306. doi: 10.1111/sji.12329. PMID: 26099424.
https://doi.org/10.1111/sji.12329 |
| 4 | Dendrou CA, Petersen J, Rossjohn J, Fugger L. HLA variation and disease. Nat Rev Immunol. 2018 May;18(5):325-339. doi: 10.1038/nri.2017.143. Epub 2018 Jan 2. PMID: 29292391.
https://doi.org/10.1038/nri.2017.143 |
| 5 | Kervevan J, Chakrabarti LA. Role of CD4+ T Cells in the Control of Viral Infections: Recent Advances and Open Questions. Int J Mol Sci. 2021 Jan 7;22(2):523. doi: 10.3390/ijms22020523. PMID: 33430234; PMCID: PMC7825705.
https://doi.org/10.3390/ijms22020523 |
| 6 | Yang P, Wang X. COVID-19: a new challenge for human beings. Cell Mol Immunol. 2020 May;17(5):555-557. doi: 10.1038/s41423-020-0407-x. Epub 2020 Mar 31. PMID: 32235915; PMCID: PMC7110263.
https://doi.org/10.1038/s41423-020-0407-x |
| 7 | Sharma AK, Sharma V, Sharma A, Pallikkuth S, Sharma AK. Current Paradigms in COVID-19 Research: Proposed Treatment Strategies, Recent Trends and Future Directions. Curr Med Chem. 2021;28(16):3173-3192. doi: 10.2174/0929867327666200711153829. PMID: 32651959.
https://doi.org/10.2174/0929867327666200711153829 |
| 8 | Melenotte C, Silvin A, Goubet AG, Lahmar I, Dubuisson A, Zumla A, Raoult D, Merad M, Gachot B, Hénon C, Solary E, Fontenay M, André F, Maeurer M, Ippolito G, Piacentini M, Wang FS, Ginhoux F, Marabelle A, Kroemer G, Derosa L, Zitvogel L. Immune responses during COVID-19 infection. Oncoimmunology. 2020 Aug 25;9(1):1807836. doi: 10.1080/2162402X.2020.1807836. PMID: 32939324; PMCID: PMC7480812.
https://doi.org/10.1080/2162402X.2020.1807836 |
| 9 | Jha SK, Imran M, Jha LA, Hasan N, Panthi VK, Paudel KR, Almalki WH, Mohammed Y, Kesharwani P. A Comprehensive Review on Pharmacokinetic Studies of Vaccines: Impact of delivery route, carrier-and its modulation on the immune response. Environ Res. 2023 Nov 1;236(Pt 2):116823. doi: 10.1016/j.envres.2023.116823. Epub 2023 Aug 3. PMID: 37543130.
https://doi.org/10.1016/j.envres.2023.116823 |
| 10 | Olvera-Collantes L, Moares N, Fernandez-Cisnal R, Muñoz-Miranda JP, Gonzalez-Garcia P, Gabucio A, Freyre-Carrillo C, Jordan-Chaves JD, Trujillo-Soto T, Rodriguez-Martinez MP, Martin-Rubio MI, Escuer E, Rodriguez-Iglesias M, Fernandez-Ponce C, Garcia-Cozar F. Development and Validation of a Highly Sensitive Multiplex Immunoassay for SARS-CoV-2 Humoral Response Monitorization: A Study of the Antibody Response in COVID-19 Patients with Different Clinical Profiles during the First and Second Waves in Cadiz, Spain. Microorganisms. 2023 Dec 16;11(12):2997. doi: 10.3390/microorganisms11122997. PMID: 38138141; PMCID: PMC10746014.
https://doi.org/10.3390/microorganisms11122997 |
| 11 | Migliorini F, Torsiello E, Spiezia F, Oliva F, Tingart M, Maffulli N. Association between HLA genotypes and COVID-19 susceptibility, severity, and progression: a comprehensive review of the literature. Eur J Med Res. 2021 Aug 3;26(1):84. doi: 10.1186/s40001-021-00563-1. PMID: 34344463; PMCID: PMC8329616.
https://doi.org/10.1186/s40001-021-00563-1 |
| 12 | Galdiero M, Galdiero M, Folliero V, Zanella C, De Filippis A, Mali A, Rinaldi L, Franci G. SARS-CoV-2 vaccine development: where are we? Eur Rev Med Pharmacol Sci. 2021 Mar;25(6):2752-2784. doi: 10.26355/eurrev_202103_25439. PMID: 33829462.
|
| 13 | Gustiananda M, Sulistyo BP, Agustriawan D, Andarini S. Immunoinformatics Analysis of SARS-CoV-2 ORF1ab Polyproteins to Identify Promiscuous and Highly Conserved T-Cell Epitopes to Formulate Vaccine for Indonesia and the World Population. Vaccines (Basel). 2021 Dec 9;9(12):1459. doi: 10.3390/vaccines9121459. PMID: 34960205; PMCID: PMC8704007.
https://doi.org/10.3390/vaccines9121459 |
| 14 | van den Dijssel J, Hagen RR, de Jongh R, Steenhuis M, Rispens T, Geerdes DM, Mok JY, Kragten AH, Duurland MC, Verstegen NJ, van Ham SM, van Esch WJ, van Gisbergen KP, Hombrink P, Ten Brinke A, van de Sandt CE. Parallel detection of SARS-CoV-2 epitopes reveals dynamic immunodominance profiles of CD8+ T memory cells in convalescent COVID-19 donors. Clin Transl Immunology. 2022 Oct 14;11(10):e1423. doi: 10.1002/cti2.1423. PMID: 36254196; PMCID: PMC9568370.
https://doi.org/10.1002/cti2.1423 |
| 15 | Jin X, Ding Y, Sun S, Wang X, Zhou Z, Liu X, Li M, Chen X, Shen A, Wu Y, Liu B, Zhang J, Li J, Yang Y, Qiu H, Shen C, He Y, Zhao G. Screening HLA-A-restricted T cell epitopes of SARS-CoV-2 and the induction of CD8+ T cell responses in HLA-A transgenic mice. Cell Mol Immunol. 2021 Dec;18(12):2588-2608. doi: 10.1038/s41423-021-00784-8. Epub 2021 Nov 2. PMID: 34728796; PMCID: PMC8561351.
https://doi.org/10.1038/s41423-021-00784-8 |
| 16 | Dijkstra JM, Frenette AP, Dixon B. Most Japanese individuals are genetically predisposed to recognize an immunogenic protein fragment shared between COVID-19 and common cold coronaviruses. F1000Res. 2021 Mar 10;10:196. doi: 10.12688/f1000research.51479.1. PMID: 34026045; PMCID: PMC8108557.
https://doi.org/10.12688/f1000research.51479.1 |
| 17 | Roche PA, Cresswell P. Antigen Processing and Presentation Mechanisms in Myeloid Cells. Microbiol Spectr. 2016 Jun;4(3):10.1128/microbiolspec.MCHD-0008-2015. doi: 10.1128/microbiolspec.MCHD-0008-2015. PMID: 27337472; PMCID: PMC6300129.
https://doi.org/10.1128/microbiolspec.MCHD-0008-2015 |
| 18 | Pan K, Chiu Y, Huang E, Chen M, Wang J, Lai I, Singh S, Shaw RM, MacCoss MJ, Yee C. Mass spectrometric identification of immunogenic SARS-CoV-2 epitopes and cognate TCRs. Proc Natl Acad Sci U S A. 2021 Nov 16;118(46):e2111815118. doi: 10.1073/pnas.2111815118. PMID: 34725257; PMCID: PMC8609653.
https://doi.org/10.1073/pnas.2111815118 |
| 19 | Mentzer AJ, O'Connor D, Bibi S, Chelysheva I, Clutterbuck EA, Demissie T, Dinesh T, Edwards NJ, Felle S, Feng S, Flaxman AL, Karp-Tatham E, Li G, Liu X, Marchevsky N, Godfrey L, Makinson R, Bull MB, Fowler J, Alamad B, Malinauskas T, Chong AY, Sanders K, Shaw RH, Voysey M; Oxford COVID Vaccine Trial Genetics Study Team Group; Snape MD, Pollard AJ, Lambe T, Knight JC. Human leukocyte antigen alleles associated with COVID-19 vaccine immunogenicity and risk of breakthrough infection. Nat Med. 2023 Jan;29(1):147-157. Doi: 10.1038/s41591-022-02078-6. Epub 2022 Oct 13. PMID: 36228659; PMCID: PMC9873562.
https://doi.org/10.1038/s41591-022-02078-6 |
| 20 | Apostolopoulos V, Bojarska J, Chai TT, Elnagdy S, Kaczmarek K, Matsoukas J, New R, Parang K, Lopez OP, Parhiz H, Perera CO, Pickholz M, Remko M, Saviano M, Skwarczynski M, Tang Y, Wolf WM, Yoshiya T, Zabrocki J, Zielenkiewicz P, AlKhazindar M, Barriga V, Kelaidonis K, Sarasia EM, Toth I. A Global Review on Short Peptides: Frontiers and Perspectives. Molecules. 2021 Jan 15;26(2):430. Doi: 10.3390/molecules26020430. PMID: 33467522; PMCID: PMC7830668.
https://doi.org/10.3390/molecules26020430 |
| 21 | Tavasolian F, Rashidi M, Hatam GR, Jeddi M, Hosseini AZ, Mosawi SH, Abdollahi E, Inman RD. HLA, Immune Response, and Susceptibility to COVID-19. Front Immunol. 2021 Jan 8;11:601886. doi: 10.3389/fimmu.2020.601886. PMID: 33488597; PMCID: PMC7820778.
https://doi.org/10.3389/fimmu.2020.601886 |
| 22 | Tambur AR, Claas FH. HLA epitopes as viewed by antibodies: what is it all about? Am J Transplant. 2015 May;15(5):1148-54. doi: 10.1111/ajt.13192. Epub 2015 Feb 23. PMID: 25708017.
https://doi.org/10.1111/ajt.13192 |
| 23 | Braun A, Rowntree LC, Huang Z, Pandey K, Thuesen N, Li C, Petersen J, Littler DR, Raji S, Nguyen THO, Jappe Lange E, Persson G, Schantz Klausen M, Kringelum J, Chung S, Croft NP, Faridi P, Ayala R, Rossjohn J, Illing PT, Scull KE, Ramarathinam S, Mifsud NA, Kedzierska K, Sørensen AB, Purcell AW. Mapping the immunopeptidome of seven SARS-CoV-2 antigens across common HLA haplotypes. Nat Commun. 2024 Aug 30;15(1):7547. doi: 10.1038/s41467-024-51959-6. PMID: 39214998; PMCID: PMC11364864.
https://doi.org/10.1038/s41467-024-51959-6 |
| 24 | Tai W, Zhang X, Drelich A, Shi J, Hsu JC, Luchsinger L, Hillyer CD, Tseng CK, Jiang S, Du L. A novel receptor-binding domain (RBD)-based mRNA vaccine against SARS-CoV-2. Cell Res. 2020 Oct;30(10):932-935. doi: 10.1038/s41422-020-0387-5. Epub 2020 Aug 5. PMID: 32759966; PMCID: PMC7403570.
https://doi.org/10.1038/s41422-020-0387-5 |
| 25 | Moss P. The T cell immune response against SARS-CoV-2. Nat Immunol. 2022 Feb;23(2):186-193. doi: 10.1038/s41590-021-01122-w. Epub 2022 Feb 1. PMID: 35105982.
https://doi.org/10.1038/s41590-021-01122-w |
| 26 | Almendro-Vázquez P, Laguna-Goya R, Paz-Artal E. Defending against SARS-CoV-2: The T cell perspective. Front Immunol. 2023 Jan 27;14:1107803. doi: 10.3389/fimmu.2023.1107803. PMID: 36776863; PMCID: PMC9911802.
https://doi.org/10.3389/fimmu.2023.1107803 |
| 27 | Slifka MK, Amanna IJ. Role of Multivalency and Antigenic Threshold in Generating Protective Antibody Responses. Front Immunol. 2019 May 1;10:956. doi: 10.3389/fimmu.2019.00956. PMID: 31118935; PMCID: PMC6504826.
https://doi.org/10.3389/fimmu.2019.00956 |
| 28 | Mettelman RC, Allen EK, Thomas PG. Mucosal immune responses to infection and vaccination in the respiratory tract. Immunity. 20 22 May 10;55(5):749-780. doi: 10.1016/j.immuni.2022.04.013. PMID: 35545027; PMCID: PMC9087965.
https://doi.org/10.1016/j.immuni.2022.04.013 |
| 29 | Yan Y, Yao D, Li X. Immunological Mechanism and Clinical Application of PAMP Adjuvants. Recent Pat Anticancer Drug Discov. 2021;16(1):30-43. doi: 10.2174/1574892816666210201114712. PMID: 33563182.
https://doi.org/10.2174/1574892816666210201114712 |
| 30 | Dan JM, Mateus J, Kato Y, Hastie KM, Yu ED, Faliti CE, Grifoni A, Ramirez SI, Haupt S, Frazier A, Nakao C, Rayaprolu V, Rawlings SA, Peters B, Krammer F, Simon V, Saphire EO, Smith DM, Weiskopf D, Sette A, Crotty S. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science. 2021 Feb 5;371(6529):eabf4063. Doi: 10.1126/science.abf4063. Epub 2021 Jan 6. PMID: 33408181; PMCID: PMC7919858.
https://doi.org/10.1126/science.abf4063 |