Original Article - DOI:10.33594/000000759
Accepted 13 February 2025 - Published online 24 February 2025
Synergistic Activity of Indolicidin and Tigecycline Against Multidrug-Resistant Acinetobacter Baumannii Clinical Isolates
Keywords
Abstract
Background/Aims:
Acinetobacter baumannii is a ubiquitous opportunistic microorganism associated with high morbidity and mortality Particularly among burn patients and immunocompromised individuals. This study aimed to assess the time-dependent antimicrobial effectiveness of indolicidin in combination with tigecycline against multidrug-resistant (MDR) A. baumannii isolates from patients with wound infections.Methods:
The antimicrobial synergy between indolicidin and tigecycline was evaluated using the checkerboard broth microdilution method And anti-biofilm activity was assessed using the crystal violet assay. Ten isolates resistant to multiple antibiotics were identified and confirmed using the API 20-NE system.Results:
Fractional inhibitory concentration (FIC) values ranged from 0.97 to 250 µg/ml. The results demonstrated that the inhibitory activity of indolicidin and tigecycline was significantly enhanced when combined With FIC values between 0.046 and 0.312 µg/ml. The MIC of indolicidin for isolate A4 decreased from 62.5 to 0.97 µg/ml • While the MIC of tigecycline decreased from 500 to 15.6 µg/ml • Indicating a strong synergistic effect. Additionally The combination therapy led to a 78% reduction in biofilm formation.Conclusion:
This study suggests a novel combination therapy for treating wound infections caused by MDR A. baumannii • Demonstrating significant efficacy in both bacterial inhibition and biofilm reduction.Introduction
Acinetobacter baumannii is a leading cause of hospital-acquired infections, particularly in ICU patients and immunocompromised individuals. Compared to other non-baumannii species, A. baumannii exhibits a high level of antibiotic resistance, leading to increased mortality rates among infected patients. Clinically, resistant strains are associated with prolonged hospital stays and higher fatality rates. This pathogen is responsible for several infections, including skin wound infections, bacteremia, and pneumonia. Inadequate treatment is one of the most common risk factors for acquiring MDR A. baumannii [1].
Bacterial infections are increasingly becoming a major public health threat due to the emergence of antibiotic resistance and the declining development of new antibiotics. As a result, preventable diseases and deaths have risen worldwide [2]. The modification and optimization of known natural antimicrobial proteins could aid in the development of new therapeutic interventions. As a class of antimicrobial peptides, antibacterial peptides exhibit broad-spectrum activity and have been extensively studied as potential treatments for bacteria resistant to multiple antibiotics [3].
Most host-defense peptides, commonly referred to as antimicrobial peptides (AMPs), are synthesized by virtually all forms of life. The mechanisms by which they function have raised significant interest. Indolicidin, an antimicrobial peptide consisting of 13 amino acids with cationic properties, was isolated from the cytoplasmic granules of bovine neutrophils. It belongs to the cathelicidin protein family. Its distinct structure differentiates it from the α-helical and β-structured cationic peptides in the current series, making it one of the smallest natural linear cationic AMPs known to date [4]. AMPs, whether used individually or in combination with other biocides, have been recommended due to their effectiveness and pharmaceutical potential [5, 6].
Multicellular organisms, including animals and plants, naturally synthesize AMPs as part of their first line of defense against pathogens. AMPs have been associated with disrupting cell membranes, weakening cell walls, inhibiting protein synthesis, altering nucleic acid dynamics, and inducing necrosis and apoptosis. The relevance of these mechanisms to A. baumannii is emphasized by its high antibiotic resistance and its ability to withstand desiccation, biocides, and ultraviolet light, making treatment particularly challenging [7, 8].
Multiple mutations in different targets within independent pathways may be necessary to overcome these challenges. An integrated approach targeting multiple pathways is essential for effectively combating MDR pathogens [9]. This study was designed to evaluate the antimicrobial synergy of indolicidin, an antimicrobial peptide, in combination with tigecycline against MDR A. baumannii strains derived from infected wound patients. Additionally, the study assessed the potential effect of this combination therapy on biofilm formation in MDR species.
Materials and Methods
Antimicrobial Peptide and Tigecycline
Indolicidin peptide (amino acid sequence ILPWKWPWWPWRR-NH2) was purchased from Northern Lipids Inc., Vancouver, BC, Canada, reconstituted in double-distilled water, and stored at -20°C until use. Tigecycline powder (Sigma-Aldrich), an antibiotic of reagent grade, was dissolved in distilled water at a concentration of 1 mg/ml and stored at -80°C until needed.
Clinical Isolates and Growth Media
Multidrug-resistant A. baumannii strains (A1 to A10) used in this study were collected from patients with wound infections at Al-Yarmouk Teaching Hospital. According to CLSI (2021) guidelines [10], organisms resistant to all agents in two or more antibiotic classes (beta-lactams, aminoglycosides, and fluoroquinolones) were classified as MDR. Species identification was confirmed using the API 20-NE system (BioMérieux, France).
Minimum Inhibitory Concentration (MIC) Determination
MIC values were determined according to CLSI (2021) guidelines. A 96-well microtiter plate was prepared by adding 180 µL of Muller-Hinton broth to the first well and 100 µL to the remaining wells. Next, 20 µL of indolicidin supernatant (500 µg/ml) was added to the first well, mixed, and serially diluted across the plate. Each well was then filled with 100 µL of a standardized bacterial solution, resulting in final indolicidin concentrations ranging from 0.97 to 250 µg/ml. Experiments were conducted in triplicate, with 100 µL of regular saline as a negative control. Plates were incubated at 37°C for 20 minutes before the addition of 20 µL resazurin dye, followed by further incubation.
Fractional Inhibitory Concentration (FIC) Determination
The antibacterial synergy of tigecycline and indolicidin was assessed using the checkerboard synergy assay and broth microdilution method. Eight MDR A. baumannii isolates were used to determine the synergistic effects of the antimicrobial agents in combination.
Bacterial strains were grown in Muller-Hinton broth and resuspended in MHB at a concentration of 1.5 × 10⁸ CFU/mL. Each antimicrobial combination was separately diluted twofold in MHB in a 96-well microtiter plate, with each well containing a final volume of 100 µL. Bacterial suspensions were then added to each well, bringing the final volume to 200 µL. Positive controls contained bacteria without antimicrobial agents, while negative controls contained only the medium. A sterile lid was placed over the plate to prevent evaporation, and the plate was shaken at 120 rpm for 18 to 24 hours at 37°C. After incubation, 20 µL of resazurin dye was added, and plates were visually inspected for sterility before further incubation for 20 minutes.
FIC was calculated using the formula:
$${ \sum \text{FICI} = \text{FIC(A)} + \text{FIC(B)} }$$
where
$${ \text{FIC(A)} = \frac{\text{MIC(A) in combination}}{\text{MIC(A) alone}} }$$
and
$${ \text{FIC(B)} = \frac{\text{MIC(B) in combination}}{\text{MIC(B) alone}} }$$
An FIC index of ≤0.5 indicated synergy, while FIC values >0.5 and ≤4.0 suggested no interaction [11].
Biofilm Formation Ability and Quantification
Biofilm formation was measured using the microtiter plate method as described by Sabo et al . [12]. A colony of A. baumannii from TSB overnight culture was adjusted to an optical density (OD600) of 0.1 (10⁸ CFU/ml) and further diluted 1:100 in fresh TSB medium. A total of 200 μL/well was inoculated into a 96-well flat-bottom microtiter plate and incubated statically at 37°C in CO₂ for 24 hours. Negative controls contained TSB medium alone.
After incubation, wells were rinsed with phosphate-buffered saline (PBS), fixed with methanol for 15 minutes, and air-dried. Wells were then stained with 0.1% crystal violet for 5 minutes, washed, and left to air dry for seven days. Glacial acetic acid (33%) was added, and absorbance was measured at 570 nm using a spectrophotometer. Biofilm formation was classified as non-biofilm producers, weak, moderate, or strong biofilm producers based on optical density values [13] as shown in Table 1.
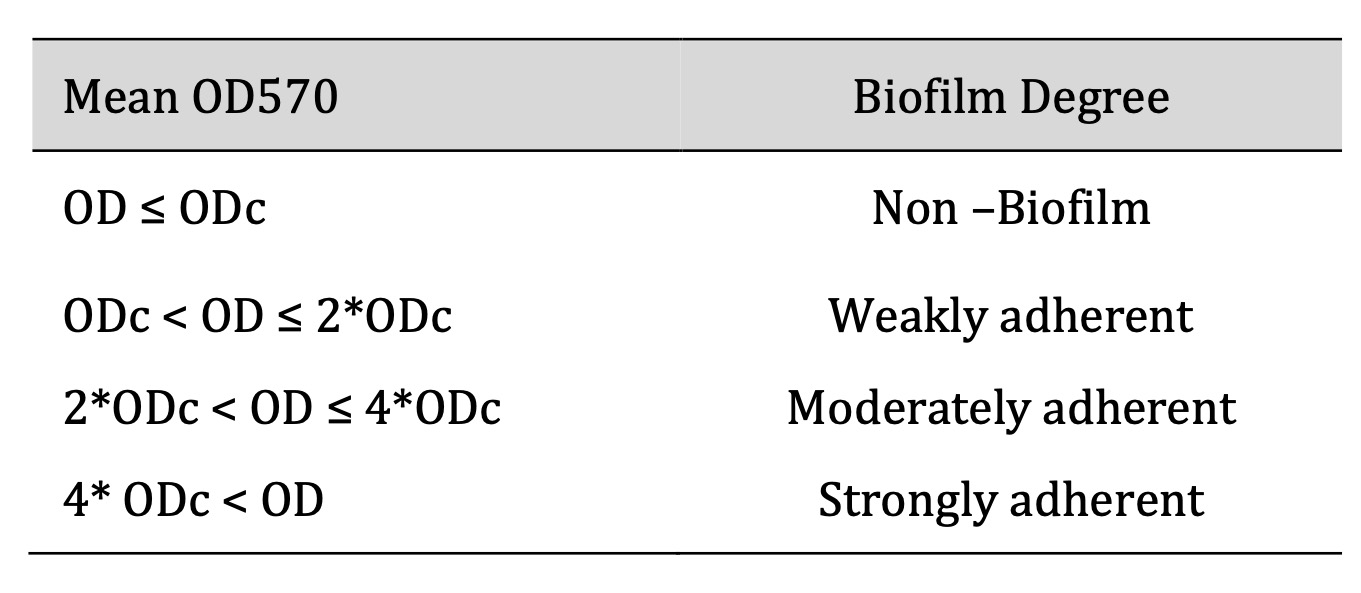
Table 1: Classification of bacterial biofilm formation. OD; optical density, ODc; cut off value (: Average OD of negative control + (3 × standard deviation (SD) of Negative control), OD isolate: Average OD of isolate – ODc
Anti-Biofilm Properties
To assess the effect of indolicidin on bacterial adherence and biofilm formation, bacterial suspensions (10⁷ CFU/ml) were mixed with peptide solutions at 1×, 1/2×, and 1/4× MIC in 96-well plates and incubated for either 2 hours (surface adhesion) or 24 hours (biofilm formation) at 37°C. After incubation, wells were washed, stained with 0.1% crystal violet, and destained with 95% ethanol. Absorbance at 570 nm was recorded using a microplate reader [14]. Statistical analysis was used for significant comparisons between values.
Results
The checkerboard assay was performed to determine the MICs of tigecycline and indolicidin and assess their interactions against A. baumannii isolates. The results are summarized in Fig. 1 and Table 2.
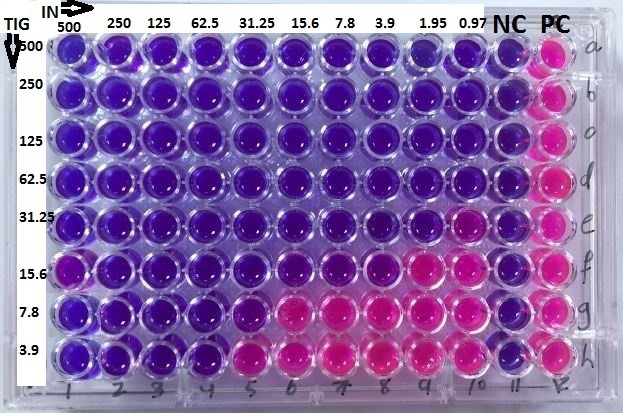
Fig. 1: Checkerboard assay results for the MIC determination of Indolicidin and Tigecycline in MDR A. baumannii isolates. NC: negative control (NC), PC: positive control (PC).
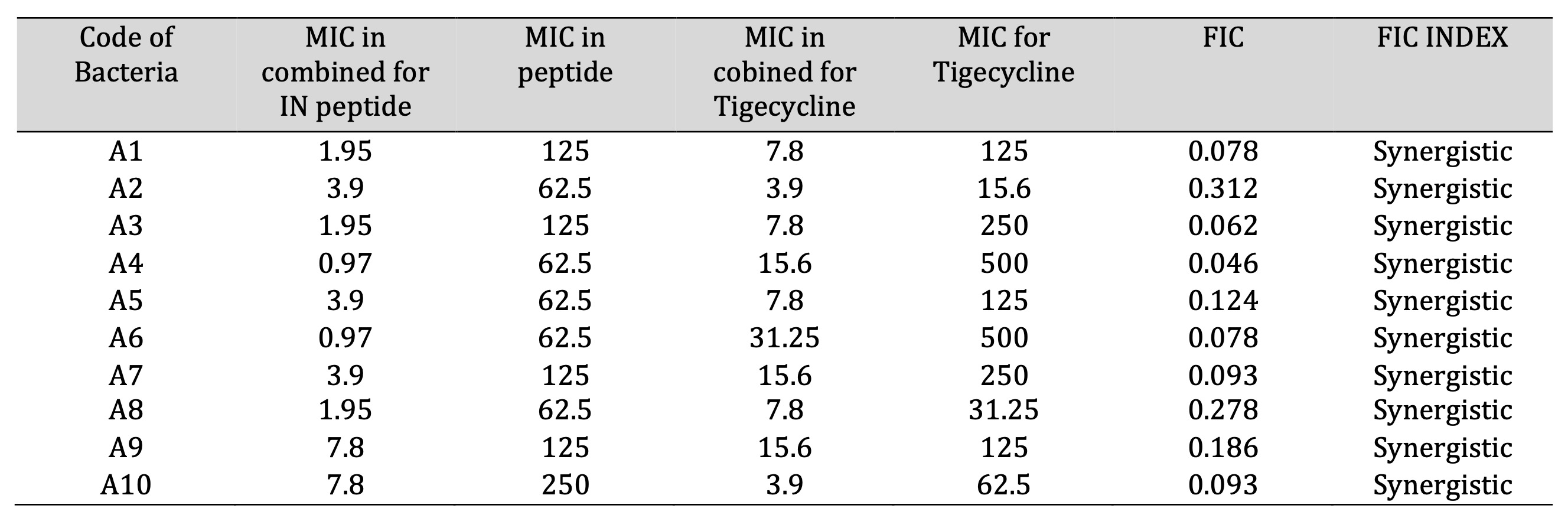
Table 2: The Fractional Inhibitory Concentrations (FICs) of Indolicidin peptide, Tigecycline against 10 A. baumannii isolates.
Indolicidin and tigecycline demonstrated reduced inhibitory capabilities when applied individually at lower concentrations, with FIC values ranging from 0.046 to 0.312 µg/ml. MIC testing across a concentration range of 0.97 to 250 µg/ml showed a synergistic effect. Specifically, for isolate A4, the MIC of indolicidin decreased from 62.5 to 0.97 µg/ml, while the MIC of tigecycline decreased from 500 to 15.6 µg/ml, demonstrating strong synergy. The FIC index results confirmed enhanced antimicrobial activity at lower concentrations.
Several studies have indicated the role of cathelicidin peptides such as LL-37 and indolicidin as effective antibacterial agents against multidrug-resistant pathogens isolated from Baghdad hospitals [15, 16, 17]. Research by Kampshoff et al. [18] showed that melimine and ciprofloxacin together exhibited synergistic efficacy against Pseudomonas aeruginosa strains with FIC values ≤0.5, regardless of antibiotic susceptibility. A proposed strategy to counter fluoroquinolone resistance in P. aeruginosa involves combining AMPs with ciprofloxacin.
At concentrations ranging from 16 to 64 mg/l, LL-37 exhibited significant activity against carbapenemase-producing E. coli strains. Checkerboard assay results further confirmed the synergy of LL-37 with other antibiotics through time-kill and post-antibiotic effect analyses [19]. Indolicidin, a cationic peptide rich in tryptophan, is known to inhibit DNA-processing enzymes and displace divalent cations, leading to membrane disruption [20].
Regarding colistin-resistant A. baumannii, MIC values ranged from 4 to 32 μg/mL, while against clinical isolates, MIC values were approximately 16 μg/mL. Indolicidin also showed activity against susceptible fungal strains, with MIC values ranging from 2 to 64 μg/mL for twelve MDR bacterial isolates [8].
Biofilm development analysis revealed that all ten A. baumannii isolates were strong biofilm producers. Treatment with indolicidin at MIC levels led to a reduction in biofilm formation ranging from 40% to 60%. The combination of indolicidin and tigecycline resulted in a more substantial reduction, as Fig. 2 shows that biofilm formation decreased by 78% compared to the control.
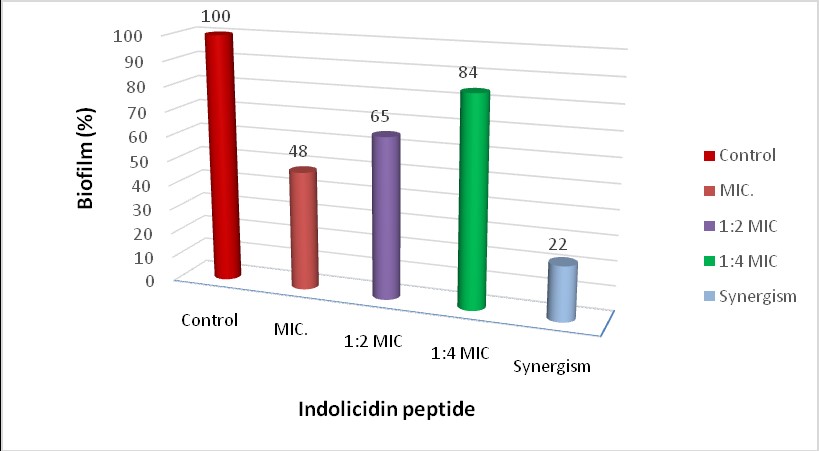
Fig. 2: Anti-biofilm activity using different concentrations) of Indolicidin peptide (1 MIC, 1/2 MIC, 1/4 MIC, and synergism with Tigecycline (0.97+15.6 µg/ml) by crystal-violet assay against A4 isolate of A. baumannii.
Discussion
Indolicidin, a 13-amino acid antimicrobial peptide, is derived from bovine neutrophil cytoplasmic granules. It exhibits broad-spectrum antimicrobial activity against Gram-positive and Gram-negative bacteria, as well as fungi. Unlike other AMPs, indolicidin interacts with bacterial membranes without causing total cell disruption. The present study confirms that indolicidin induces morphological changes in Escherichia coli, leading to filamentation [22].
AMP-antibiotic combinations have demonstrated significant antimicrobial synergy. Nisin, when used with antibiotics such as penicillin, chloramphenicol, ciprofloxacin, indolicidin, or azithromycin, exhibited an additive or synergistic effect in biofilm inhibition, preventing bacterial adherence to surfaces and impeding the growth of methicillin-resistant Staphylococcus aureus [23].
The antimicrobial efficacy of AMPs against biofilms is attributed to multiple mechanisms. These include altering electrochemical properties of bacterial cells within biofilms, impairing bacterial communication pathways, degrading biofilm matrices and polysaccharides, inhibiting the bacterial stringent response, and suppressing biofilm-regulating genes responsible for electron transport and protein binding [24].
This study highlights the potential of indolicidin as an effective antimicrobial peptide against MDR A. baumannii, especially in combination with tigecycline. The significant reduction in MIC values and biofilm formation suggests that this combination could serve as an alternative therapeutic strategy against MDR infections. Further studies, including in vivo models and clinical trials, are recommended to validate these findings and assess the safety and efficacy of the combination therapy.
Conclusion
This study demonstrated the potent antimicrobial synergy between indolicidin and tigecycline against MDR A. baumannii. The combination therapy significantly reduced bacterial growth and biofilm formation, suggesting a novel approach for treating wound infections caused by MDR A. baumannii.
Disclosure Statement
The auhtors have nothing to disclose.
AI Disclosure
No AI tools have been used to create this work.
References
| 1 | Reina R, León-Moya C, Garnacho-Montero J.: Treatment of Acinetobacter baumannii severe infections. Med Intensiva 2022;46:700-710.
https://doi.org/10.1016/j.medin.2022.08.003 |
| 2 | Ventola, C.L.: The antibiotic resistance crisis. Pharm Ther 2015;40:277-283.
|
| 3 | Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q.: The antimicrobial peptides and their potential clinical applications. Am J Transl Res 2019;11:3919-3931.
|
| 4 | Bucataru C, Ciobanasu C.: Antimicrobial peptides: Opportunities and challenges in overcoming resistance. Microbiol Res 2024;286:127822.
https://doi.org/10.1016/j.micres.2024.127822 |
| 5 | Naveed, M., Naveed, R., Aziz, T., Iqbal, F., Hassan, A., Saleem, A., Waseem M., Rahman S. Alharbi M., Alshammari A., Alasmari, F., : Exploring the potential application of peroxidase enzyme from Acinetobacter baumannii as an eco-friendly agent for the bioremediation of the highly noxious pyrethroid compounds through molecular docking analysis. Biomass Conversion and Biorefinery 2023;15:155-173.
https://doi.org/10.1007/s13399-023-05160-2 |
| 6 | Naveed, M., Abid, A., Aziz, T., Saleem, A., Arshad, A., Javed, K., Rehman, H. M., Nabi, G., Al-Harbi, M., & Alasmari, A. F.: Fragment-optimized chalcone derivatives targeting OmpA protein as a therapeutic approach against multidrug-resistant Acinetobacter baumannii. Sci Rep 2025;15:3917.
https://doi.org/10.1038/s41598-025-88191-1 |
| 7 | Mhlongo JT, Waddad AY, Albericio F, de la Torre BG.: Antimicrobial Peptide Synergies for Fighting Infectious Diseases. Adv Sci (Weinh) 2023;10:e2300472.
https://doi.org/10.1002/advs.202300472 |
| 8 | Rangel K, Lechuga GC, Provance DW Jr, Morel CM, De Simone SG.: An Update on the Therapeutic Potential of Antimicrobial Peptides against Acinetobacter baumannii Infections. Pharmaceuticals 2023;16:1281.
https://doi.org/10.3390/ph16091281 |
| 9 | Duong, L.; Gross, S.P.; Siryaporn, A.: Developing Antimicrobial Synergy with AMPs. Front Med Technol 2021;3:640981.
https://doi.org/10.3389/fmedt.2021.640981 |
| 10 | Clinical Laboratory Standards Institute (CLSI). (2021). Performance standards for antimicrobial susceptibility testing; 21st informational supplement, CLSI M100-S21, Clinical and Laboratory Standards Institute Wayne, PA, U.S.A.
|
| 11 | Odds, F.C.:Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother 2003;52:1.
https://doi.org/10.1093/jac/dkg301 |
| 12 | Sabo, I. P., Bjoras, M., Franzyk, H., Helgesen, E. & Booth, J. A. (2023). Optimization of the hemolysis assay for the assessment of cytotoxicity. Int. J. Mol. Sci. 2, 2914.
https://doi.org/10.3390/ijms24032914 |
| 13 | Bezek, K., Avberšek, J., Zorman-Rojs, O. & Barlič-Maganja, D.:. Antimicrobial and antibiofilm effect of commonly used disinfectants on Salmonella infantis isolates. Microorganism 2023;11:301.
https://doi.org/10.3390/microorganisms11020301 |
| 14 | Mataraci, E., Mataraci, D. S. & Dosler, S.: In vitro activities of antibiotics and antimicrobial cationic peptides alone and in combination against methicillin-resistant Staphylococcus aureus biofilms. Antimicrob. Agent. Chemother 2012;56:49.
https://doi.org/10.1128/AAC.01180-12 |
| 15 | AL-Sabagh, F. S. H. and Ghaima, K. K.:The antibacterial activity of LL-37 peptide against multidrug-resistant Pseudomonas aeruginosa isolated from burn infections. Revista Bionatura 2023;8:69.
https://doi.org/10.21931/RB/2023.08.01.69 |
| 16 | Zaidan, N.I., Ghaima, K.K., Al-Haboobi, H.M.R.:Antibacterial Activity of Antimicrobial Peptide Indolicidin against Multidrug-Resistant Klebseilla pneumoniae Isolated from Patients with Burns. Indian Journal of Forensic Medicine & Toxicology 2022;16:282-290.
https://doi.org/10.37506/ijfmt.v16i2.17973 |
| 17 | Al-Sabagh, F.S., Kais K. Ghaima, K.K.: Synergistic Effect of Antimicrobial Peptide LL-37 and Ciprofloxacin against Multidrug Resistant Pseudomonas aeruginosa Isolated from Burn Infections. Iraqi journal of biotechnology 2022;21:32-38.
|
| 18 | Kampshoff, F.; Willcox, M.D.P.; Dutta, D. A.: Pilot Study of the Synergy between Two Antimicrobial Peptides and Two Common Antibiotics. Antibiotics 2019;8:60.
https://doi.org/10.3390/antibiotics8020060 |
| 19 | Morroni, G.; Sante, L.D.; Simonetti, O.; Brescini, L.,; Kamysz, W.; Kamysz, E.; Mingoia, M.; Brenciani, A.; Giovanetti, E.; Bagnarelli, P.; Giacometti, A.; Cirioni, O.: Synergistic effect of antimicrobial peptide LL-37 and colistin combination against multidrug-resistant Escherichia coli isolates. Future Microbiol 2021;16,221-227.
https://doi.org/10.2217/fmb-2020-0204 |
| 20 | Marchand C., Krajewski K., Lee H.F., Antony S., Johnson A.A., Amin R., Kvaratskhelia M., Pommier Y.: Covalent binding of the natural antimicrobial peptide indolicidin to DNA abasic sites. Nucleic Acids Res 2006;34:5157-5165.
https://doi.org/10.1093/nar/gkl667 |
| 21 | Vila-Farres X., De La Maria C.G., López-Rojas R., Pachón J., Giralt E., Vila J.: In vitro activity of several antimicrobial peptides against colistin-susceptible and colistin-resistant Acinetobacter baumannii. Clin Microbiol Infect 2012;18:383-387.
https://doi.org/10.1111/j.1469-0691.2011.03581.x |
| 22 | Subbalakshmi C, Sitaram N.: Mechanism of antimicrobial action of indolicidin. FEMS Microbiol Lett 1998;1;160:91-96.
https://doi.org/10.1111/j.1574-6968.1998.tb12896.x |
| 23 | Zhang QY, Yan ZB, Meng YM, Hong XY, Shao G, Ma JJ, Cheng XR, Liu J, Kang J, Fu CY.: Antimicrobial peptides: mechanism of action, activity and clinical potential. Mil Med Res 2021;9;8:48.
https://doi.org/10.1186/s40779-021-00343-2 |
| 24 | Yasir, M.; Willcox, M.D.P.; Dutta, D. Action of Antimicrobial Peptides against Bacterial Biofilms. Materials 2018;11:2468.
https://doi.org/10.3390/ma11122468 |