Original Article - DOI:10.33594/000000729
Accepted 6 September 2024 - Published online 23 September 2024
Peroxyredoxin 6 Protects RIN-M5F Pancreatic Beta Cells Against Streptozotocin-Induced Senescence
Keywords
Abstract
Background/Aims:
There are evidences that a decrease in the functional activity of pancreatic β-cells under type 2 diabetes conditions may be associated with their senescence, therefore, senotherapy may be a prospective strategy for the diabetes treatment.Methods:
The senotherapeutic potential of peroxiredoxin 6 (PRDX6) was studied in RIN-m5F pancreatic β-cells with streptozotocin-induced senescence by measuring markers, associated with senescence.Results:
Exposure to streptozotocin (STZ) resulted in the senescence of the β-cells. The addition of PRDX6 to the culture medium of RIN-m5F β-cells before treatment with STZ decreased the levels of the following senescence markers: the percentage of SA-β-Gal-positive cells, the phosphorylation of histone H2AX and p21 proteins, and the secretion of the proinflammatory cytokine IL-6 but not the anti-inflammatory cytokine IL-10. These effects were accompanied by a decrease in the production of reactive oxygen species (ROS) and the restoration of impaired NF-κB activation. In addition, PRDX6 altered the production of the heat shock protein HSP90: the production of the constitutive form of HSP90-beta decreased, while the level of inducible HSP90-alpha increased.Conclusion:
PRDX6 prevented the senescence of RIN-m5F cells in response to the DNA damage-inducing agent streptozotocin, indicating a potential protective role of PRDX6 in type 2 diabetes mellitus.Introduction
Epidemiological studies of diabetes mellitus are regularly conducted by the World Health Organization (WHO) and the International Diabetes Federation (IDF). In 1998, researchers predicted that the number of patients with type 1 and type 2 diabetes would increase from 135 million in 1995 to 300 million by 2025 [1]. As of the end of 2021, 537 million patients were suffering from type 1 and 2 diabetes, and according to forecasts, this number will reach 783 million in 2045 [2]. The present study aimed to address an important medical and biological problem associated with treating type 2 diabetes mellitus (T2D). Among all of the cases of diabetes mellitus that are diagnosed every year, more than 90% of patients suffer from type 2 diabetes [3, 4]. A main symptom of both types of diabetes is an increase in the concentration of glucose in the blood. Although type 1 diabetes (T1D) is insulin-dependent, associated with the destruction of pancreatic β-cells, and patients must take insulin for their whole life, T2D patients do not require lifelong insulin administration. Furthermore, their pancreatic β-cells only partially lose their functional activity [5], as shown by the loss of their secretory function and a decrease in β-cell mass [6].
Type 1 diabetes (T1D), which accounts for approximately 5% of all diabetes cases [7], occurs when β cells are selectively destroyed by an autoimmune process. The development of autoimmune diabetes is preceded by the infiltration of pancreatic islets by immune cells. Ultimately, the breakdown of self-antigen tolerance allows autoreactive T cells to become activated and attack β-cells, resulting in a loss of insulin secretion. However, innate immune cells, such as macrophages and dendritic cells (DCs), are the first cells to invade islets during insulitis [8–10].
More than three decades ago, it was shown that glutathione peroxidase activity and peroxide resistance are approximately 20 times greater in the liver and kidneys than in the pancreas [11]. This important finding indicates the natural vulnerability of pancreatic β-cells to oxidative stress. Hyperglycemia leads to the production of reactive oxygen species (ROS) and reduces the effectiveness of endogenous antioxidant defense systems in diabetes mellitus [12]. The antioxidant defense mechanisms include both enzymatic and nonenzymatic systems. There is convincing evidence of oxidative stress in both type 1 and type 2 diabetes, which is accompanied by the death and senescence of β-cells [13]. Furthermore, increased production of reactive oxygen or nitrogen species may deplete endogenous antioxidant enzymes (superoxide dismutase, glutathione peroxidase and catalase) and decrease the concentration of some low molecular weight antioxidants, such as vitamin D [14]. A decrease in the activity of peroxiredoxins under conditions of oxidative and carbonyl stress may be an important factor triggering the molecular mechanisms of oxidative damage in the vascular wall in atherosclerosis and diabetes mellitus [15, 16]. Accounting for the reduced activity of endogenous antioxidant enzymes, more attention has been given to the correction of pathology by the use of exogenous administration of natural protein preparations, such as PRDX6, which has antioxidant activity and can protect pancreatic β-cells during the development of diabetes.
We believe that the main reason for the decrease in the functional activity of pancreatic β-cells under T2D conditions may be associated with their aging or senescence. Oxidative stress is often accompanied by DNA damage, leading to a so-called DNA damage response, manifested as proliferation arrest and the expression of senescence markers. These cells do not die but partially retain their functions, though they are dysregulated in some ways. Additionally, these cells acquire a senescence-associated secretory phenotype (SASP), which is characterized by the production of proinflammatory mediators, among others, that aggravate oxidative stress. We propose that such a shift in β-cells may account for some features of type 2 diabetes.
In the present study, we used a T2D cell model. This involved culturing rat insulinoma β-cells (RIN-m5F) in the presence of streptozotocin (STZ), a known inducer of both insulin-dependent and non-insulin-dependent diabetes mellitus in animals. STZ selectively affects pancreatic β-cells through the glucose transporter GLUT2, triggering DNA alkylation and the generation of reactive oxygen and nitrogen species, leading to changes in pancreatic β-cell DNA, including DNA fragmentation. STZ-induced DNA damage activates poly-ADP ribosylation. This process leads to the depletion of cellular NAD+, a further decrease in ATP content and subsequent inhibition of insulin synthesis and secretion [17] and provokes cell senescence [18].
Previously, we showed that the natural antioxidant protein PRDX6 has a wide range of biological activities, one of which is a pronounced senomorphic effect. When administered exogenously, this protein can neutralize ROS during genotoxic stress caused by ionizing radiation, both in the extracellular environment and inside the cell. This is due to its ability to undergo intracellular translocation resulting from phospholipase activity and modulation of TLR4/NF-κB signaling [19, 20]. We were the first to show that PRDX6 is an agonist of the TLR4 receptor complex [19], which regulates insulin homeostasis and β-cell viability [21]. In this vein, PRDX6 appears to be an attractive agent in senotherapy for antidiabetic therapy.
Notably, we studied the mechanisms underlying the senoprotective effect of exogenous PRDX6 using 3T3 fibroblasts. Moreover, we demonstrated that PRDX6 penetrates 3T3 fibroblasts, which is important for potential biomedical applications [22]. More recent work has assessed the role of peroxiredoxin 6 in neutralizing X-ray-induced oxidative stress and its effect on gene expression, the preservation of radiosensitive tissues and postirradiation survival in animals [23]. In the mentioned work, the effective dose of PRDX6 was assessed.
In the present work, the mechanisms by which PRDX6 affects cell senescence were investigated via the accumulation of senescence markers (SA-β-Gal, H2AX, and p21). In addition, the effect of PRDX6 on inflammatory and stress response levels was assessed by measuring the synthesis of proinflammatory cytokines; the transcription factor NF-κB; and the heat shock proteins HSP70, HSP90β and HSP90α in RIN-m5F cells. PRDX6 may exert a senomorphic effect on RIN-m5F β-cells, indicating the possibility of reducing the pathological consequences of type 2 diabetes mellitus.
Materials and Methods
Culture of RIN-m5F β-cells
RIN-m5F
cells were grown in culture flasks in a medium consisting of a
mixture of RPMI/DMEM (PanEco, Russia) at a 1:1 ratio (total glucose
content 8.0 mM) supplemented with 10% fetal calf serum and a mixture
of antibiotics and antimycotics (Merck, Germany) in accordance with
the manufacturer’s recommendations at 37 °C and 5% CO2.
Mycoplasma testing has been carried out for the cell line. After 3-7
passages, the cells were used in the experiments. Streptozotocin
(Sigma‒Aldrich, USA) was added to the culture medium at a final
concentration of 4 mM. Peroxiredoxin 6 was obtained using genetically
engineered constructs that were subsequently used to transform E.
coli BL21 (DE3) cells [24]. Peroxiredoxin 6 at a concentration of
150 μg/ml was added together with or 1 hour before STZ. Within each
independent experiment, the characteristics of the samples were
measured in parallel in 9-12 repetitions, and an average value was
obtained. The average values from four experiments were used to
determine the significance of differences between groups (n=4). Cells
that were not subjected to additional treatment with STZ and/or PRDX6
were used as controls.
Measurement of oxidative stress in RIN-m5F β-cells
The
level of oxidative stress was measured using the diacetyl derivative
of 5, 6-carboxy-2',7'-dichlorodihydrofluorescein (carboxy-H2DCFDA)
(Invitrogen, USA). Carboxy-H2DCFDA is a chemical analog of
fluorescein and is commonly used as an indicator of oxidative stress
in cells. This nonfluorescent molecule is readily converted to the
green fluorescent molecule form (carboxy-DCF) after the intracellular
removal of acetate groups by intracellular esterases. DCFH (and
carboxy-H2DCFDA) do not react with superoxide, hydrogen peroxide, or
nitric oxide. The DCFH/carboxy-H2DCFDA ratio reflects
peroxidase-catalyzed oxidation. RIN-m5F cells were precultured for 24
h in a 96-well plate (2.5 × 104 cells per well in 100 μl
of DMEM), washed with PBS, and filled with a freshly prepared
solution of carboxy-H2DCFDA (Invitrogen, USA) in sterile DMSO at a
concentration of 2.5 μM in DMEM supplemented with 2% FTS. RIN-m5F
cells were incubated with carboxy-H2DCFDA in the dark for 1 hour.
With the addition of carboxy-H2DCFDA, other reagents were added.
Cells treated with carboxy-H2DCFDA without other agents were used as
controls. Fluorescence was measured using an Infinite 200
fluorescence plate reader (Tecan, Austria) at an excitation
wavelength of 480 nm and an absorption wavelength of 530 nm (Ex/Em =
485/535 nm). Three independent experiments were performed (three
repetitions for each experimental group), with nine technical
replicates in each group (n = 9).
Measurement of survival (cytotoxicity test)
RIN-m5F
cells (2x104 cells in 0.1 ml per well) were cultured in
96-well plates in RPMI 1640 medium supplemented with 10% fetal calf
serum, 0.2% L-glutamine and a mixture of antibiotics and antimycotics
at 37 °C and 5% CO2. After 24 hours of incubation, 1
μg/ml actinomycin D (Sigma‒Aldrich, USA) was added to the formed
monolayer of RIN-m5F β-cells, and PRDX6 and/or STZ were then added.
After 24 hours of incubation, the washed monolayer was stained for 10
minutes with a 0.05% solution of crystal violet (Sigma‒Aldrich,
USA), the wells were thoroughly washed with tap water, and 100 μl of
1% sodium dodecyl sulfate was added to each well. After 10 min, the
optical density was measured at 546 nm using a plate
spectrophotometer (Multiscan EX; Thermo Electron Corporation, USA).
Cell viability (%) was calculated using control samples as a
reference.
Cytokine production measurement
Twenty-four
hours after the addition of STZ and/or peroxiredoxin 6, the RIN-m5F
cells were removed from the 24-well plates and centrifuged for 5 min
at 2000 rpm, after which the supernatants were collected and used for
immediate analysis. For ELISA, we used primary rabbit antibodies
against mouse IL-6, IL-10, or TNFα (all from Affinity Biosciences,
China); secondary goat antibodies against rabbit immunoglobulins IgG,
IgA, and IgM conjugated with biotin (Imtek, Russia); and horseradish
peroxidase conjugated with staphylococcal streptavidin (Imtek,
Russia). The cells were incubated with antibodies according to the
manufacturer's recommendations. Each stage of the ELISA was
accompanied by washing the wells three times with a PBS solution
containing 0.05% Tween 20. To visualize binding, the TMB substrate
for horseradish peroxidase was used in an aqueous solution of 0.1 M
sodium citrate (pH 5.4) containing a source of hydrogen peroxide
(Imtek, Russia), and the reaction was stopped by adding an equal
volume of 5% sulfuric acid. Optical density was measured at 450 nm
using a microplate spectrophotometer (Multiscan EX; Thermo Electron
Corporation, USA).
Assessment of the senescent RIN-m5F cell count
The
senotherapeutic effect of the preliminary application of PRDX6 was
studied by adding PRDX6 to the cell culture medium 1 hour before the
addition of STZ. Additionally, the protective effect of the
combination of STZ and PRDX6 was studied. The cells were cultured for
120 hours at 37 °C and 5% CO2 and then fixed, and
SA-β-Gal activity was determined by the intensity of X-gal staining,
as described previously [25]. The samples were examined and
photographed using an Axio Imager M1 light scanning microscope (Carl
Zeiss, Germany). The number of senescent cells was assessed by the
percentage of SA-β-Gal-positive cells using the ImageJ program (NIH,
USA).
SDS‒PAGE electrophoresis and immunoblotting
To
prepare protein samples from RIN-m5F β-cells, the cell suspension
was washed out of the culture medium with 0.9% (w/v) NaCl and
subjected to three freeze‒thaw cycles. Before the freeze‒thaw
cycles, protease and phosphatase inhibitors (100×) were added to the
suspension according to the manufacturer’s recommendations (Thermo
Fisher Scientific, USA). Solubilization buffer for electrophoresis
and β-mercaptoethanol (2–5%) were then added, and the samples were
boiled for 5 min. The protein concentration was measured using a
NanoDrop2000 spectrophotometer (NanoDrop, USA). Protein samples (10
μl) were added to each well for 10% (w/v) Ds-Na-PAGE. The presence
of proteins in the samples was determined by Western blot analysis
using the following sets of primary antibodies: rabbit antibodies
against phH2AX (AF3187, Affinity, China), p21 (AF6290, Affinity,
China), HSP90α (BF0084, Affinity, China), HSP90β (BF0215, Affinity,
China), total NF-κB (total-NF-κB) (AF5006, Affinity, China), NF-κB
phosphorylated at Ser536 (ph-NF-κB (Ser536)) (AF2006, Affinity,
China), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH)
(PAB932Mu01, Cloud-Clone Corp, China). Goat antibodies conjugated to
rabbit immunoglobulins IgG, IgA, and IgM with horseradish peroxidase
(P-GAR, Imtek, Russia) were used as secondary antibodies. An enhanced
chemiluminescence (ECL) system (Bio-Rad, Germany) was used to
identify proteins. Images of the blots were acquired using a TFX
35.WL transilluminator (Vilber Lourmat, France). Proteins were
quantified by densitometry using the Qapa program (ver. 3.7, Russia).
Three independent experiments were carried out (cells from different
passages were used) for each protein. The resulting digital data were
normalized to the corresponding loading control (GAPDH bands) and are
expressed in relative units.
Statistical analysis
Statistical
analysis was performed using the Statistica 6.0 software package
(StatSoft, USA). To determine the significance of differences,
one-way analysis of variance (ANOVA) with Tukey's post hoc test was
used. Differences were considered significant at p < 0.05. The
Shapiro–Wilk test was used to check the normality of the
distribution. The W value was not significant in any case (p >
0.05).
Results
PRDX6 reduces STZ-induced senescence of RIN-m5F cells
Streptozotocin
is particularly toxic to insulin-producing β-cells of the mammalian
pancreas. The genotoxic effect of this agent led to increased
senescence of these cells. Our data showed that the presence of STZ
in the RIN-m5F culture medium led to an increase in the number of
senescent cells in the culture (Fig. 1a, 1b) and the accumulation of
the senescence markers ph-H2AX and p21 (Fig. 2).
In
the presence of PRDX6, the percentage of SA-β-Gal-positive cells
decreased (Fig. 1b), as did the levels of the senescence marker p21
protein and the phosphorylated form of histone H2AX (Fig. 2),
provided that PRDX6 was added to the culture medium 1 h before STZ
treatment.
Our
results indicated that PRDX6 has a senescence-preventive effect on
RIN-m5F rat insulinoma β-cells. The preliminary addition of PRDX6
prevented STZ-induced senescence of β-cells, as assessed by the
number of SA-β-Gal-positive cells, and the levels of the senescence
markers p21 and phH2AX also decreased. This finding indicates a
pronounced senotherapeutic effect of the antioxidant protein PRDX6
and confirms the senomorphic activity of this protein that we
previously discovered (22, 26) in relation to the pathological
consequences of T1D.
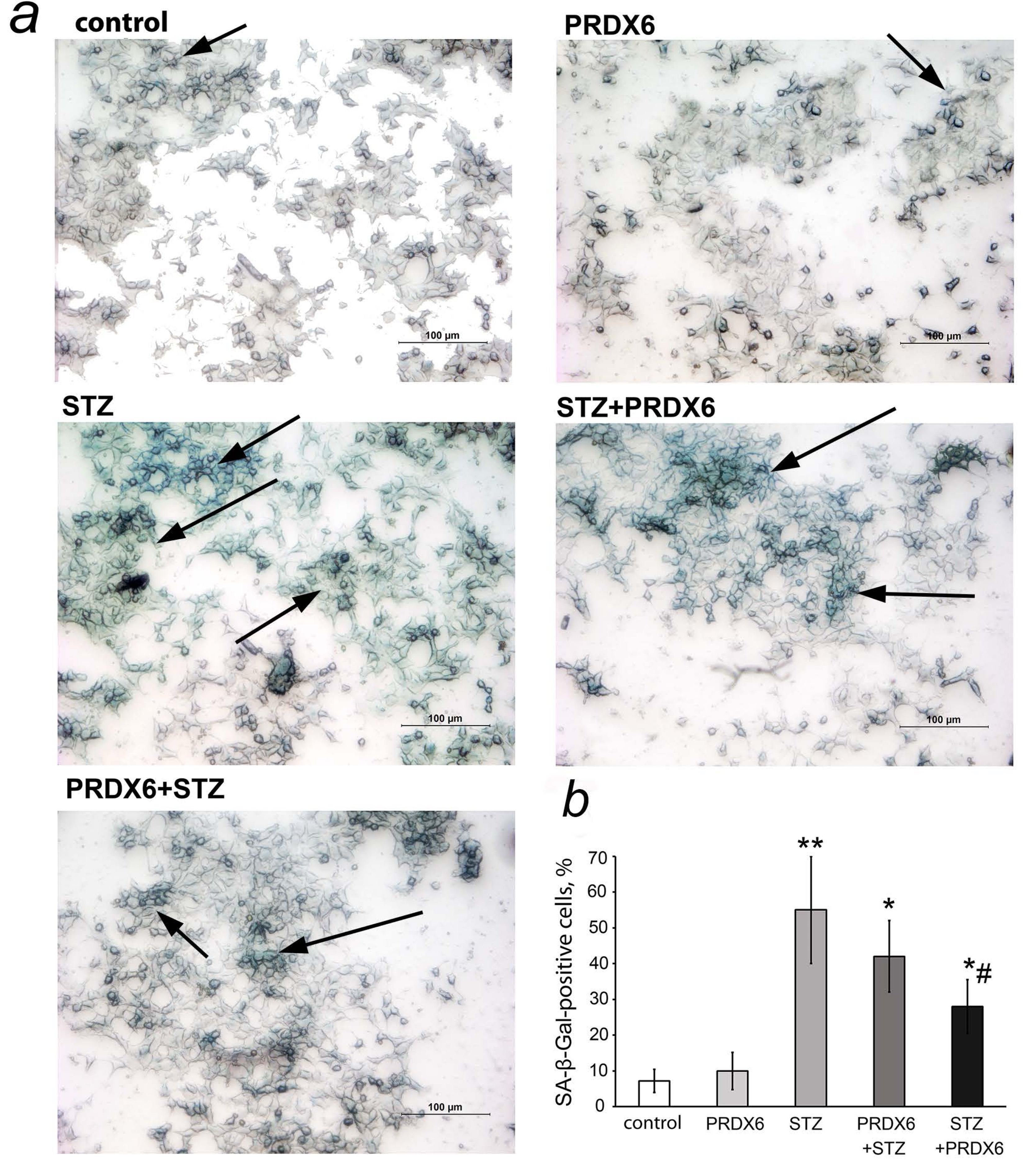
Fig. 1: Effect of streptozotocin and peroxiredoxin 6 on the accumulation of the senescence marker SA-β-Gal in RIN-m5F cells. Panel a: SA-β-Gal-positive cells (green staining) in groups without drug treatment (control), 150 μg/ml peroxiredoxin 6 (PRDX6), 4 mM streptozotocin (STZ), peroxiredoxin 6 plus streptozotocin simultaneously (STZ +PRDX6) and peroxiredoxin 6 one hour before streptozotocin (PRDX6+STZ). Arrows on microphotographs indicate groups of stained senescent cells. Panel b: Percentage of SA-β-Gal-positive cells of the total number of cells in the field of view according to image processing data. Counting was carried out in 4-5 fields of view, and the average value and standard deviation were calculated. *- significant difference from the control group (p<0.05), **- significant difference from the control group (p<0.01), # - significant difference from the STZ group (p<0.05)
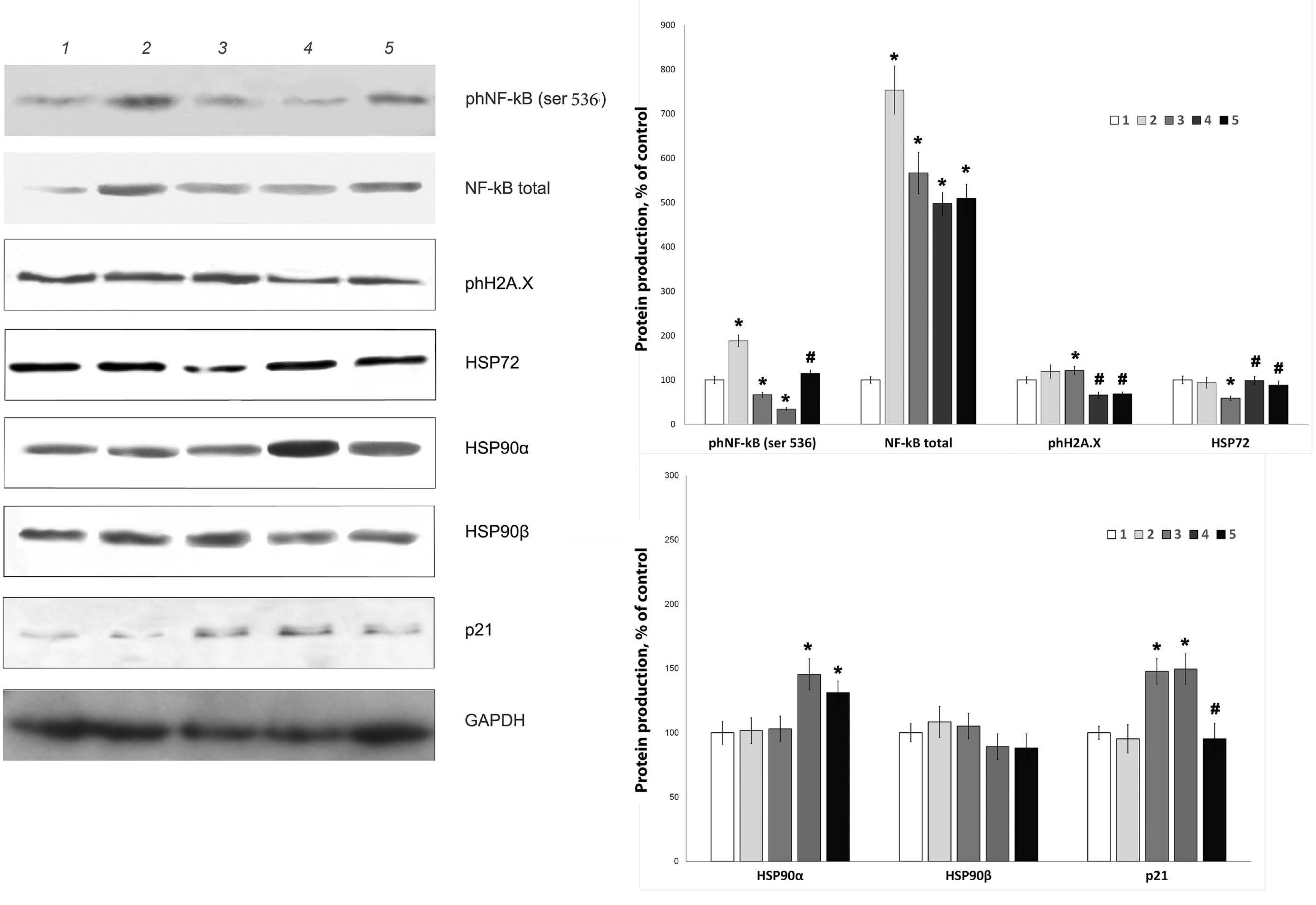
Fig. 2: Effect of streptozotocin and peroxiredoxin 6 on the production of senescence-related proteins in RIN-m5F cells. Representative images of protein bands obtained by Western blotting and densitometry are shown. 1 - control, 2 - PRDX6, 3 - STZ, 4 - STZ+PRDX6 simultaneously, 5 - PRDX6 one hour before STZ. The values are the means of 3 independent experiments ± SEMs normalized to the protein loading control (GAPDH) and to the control group. * - significant difference from the control group (p<0.05), # - significant difference from the STZ group (p<0.05)
Peroxiredoxin 6 normalizes the oxidative stress level and proinflammatory cytokine production in streptozotocin-treated RIN-m5F β-cells
Our
data showed that the addition of STZ to the culture medium of RIN-m5F
β-cells stimulated the production of ROS and proinflammatory
cytokines, supporting the oxidative stress-inducing effect of STZ in
β-cells (Figs. 3, 4). The main sign of the protective effect of
antioxidants is a decrease in the production of ROS in cells, and
this decrease was demonstrated in our experiments (Fig. 3).
STZ
increased the concentration of the proinflammatory cytokine IL-6 but
not the anti-inflammatory cytokine IL-10. PRDX6 decreased the level
of the proinflammatory cytokine IL-6 but not the anti-inflammatory
cytokine IL-10 (Fig. 4).
However,
when PRDX6 was combined with STZ, the protective effect of the
antioxidant was less significant than when it was added to the
culture medium 1 hour before STZ, but this effect was only observed
for the proinflammatory cytokine IL-6. The activation of the
anti-inflammatory cytokine IL-10 increased with the simultaneous
addition of STZ and PRDX6 to the cell culture medium and remained
significantly increased in the presence of PRDX6 compared with that
in the control group.
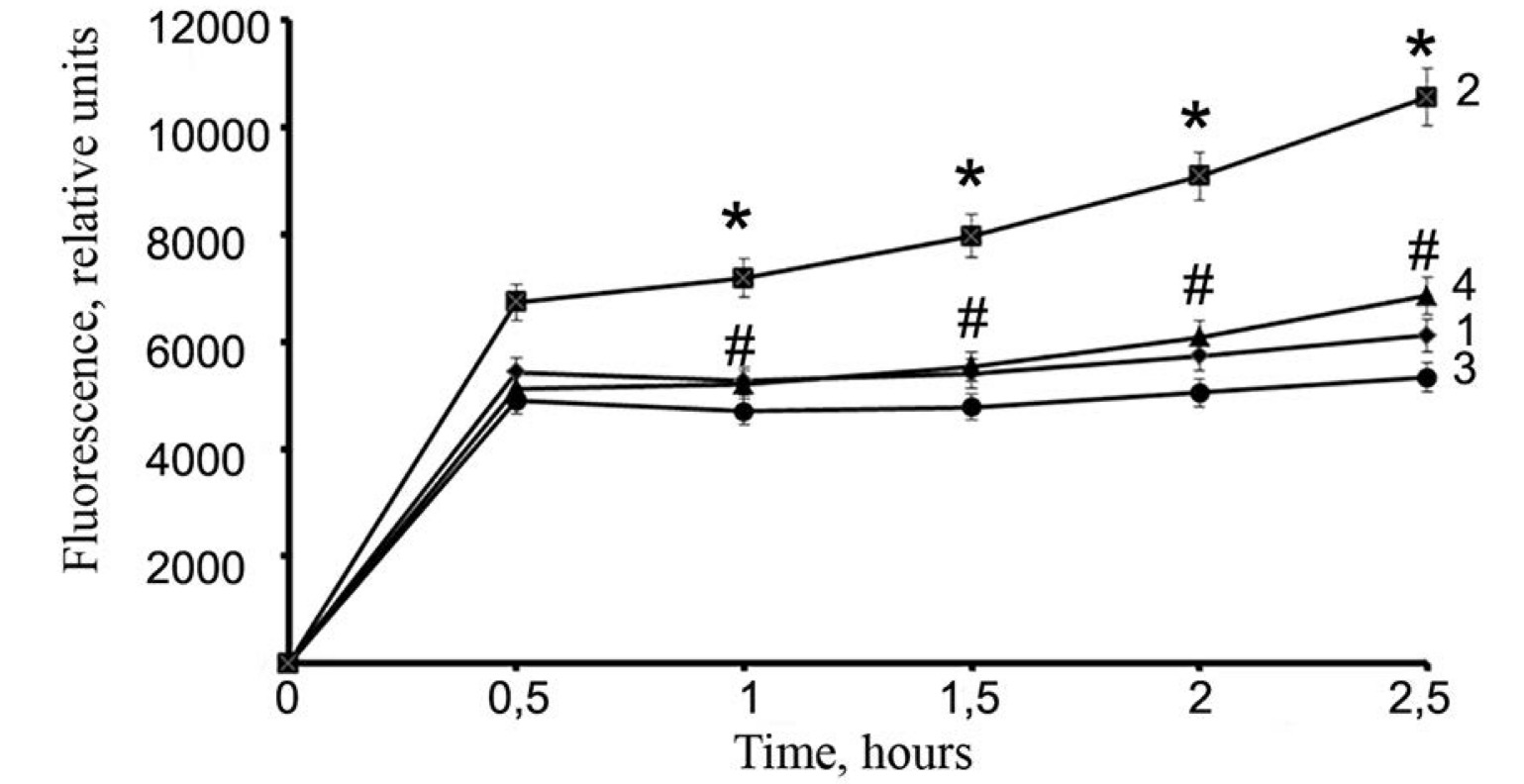
Fig. 3: Effects of streptozotocin and peroxiredoxin 6 on ROS production in RIN-m5F cells. Changes in the fluorescence intensity of the carboxy-H2DCFDA probe were observed at an excitation wavelength of 480 nm and an absorption wavelength of 530 nm upon the addition of STZ and PRDX6. 1 - control, 2 - STZ, 3 - PRDX6, 4 - PRDX6 and STZ. The values are the means of 3 independent experiments ± SEMs. * - significant difference from the control group (p<0.05), # - significant difference from the STZ group (p<0.05)
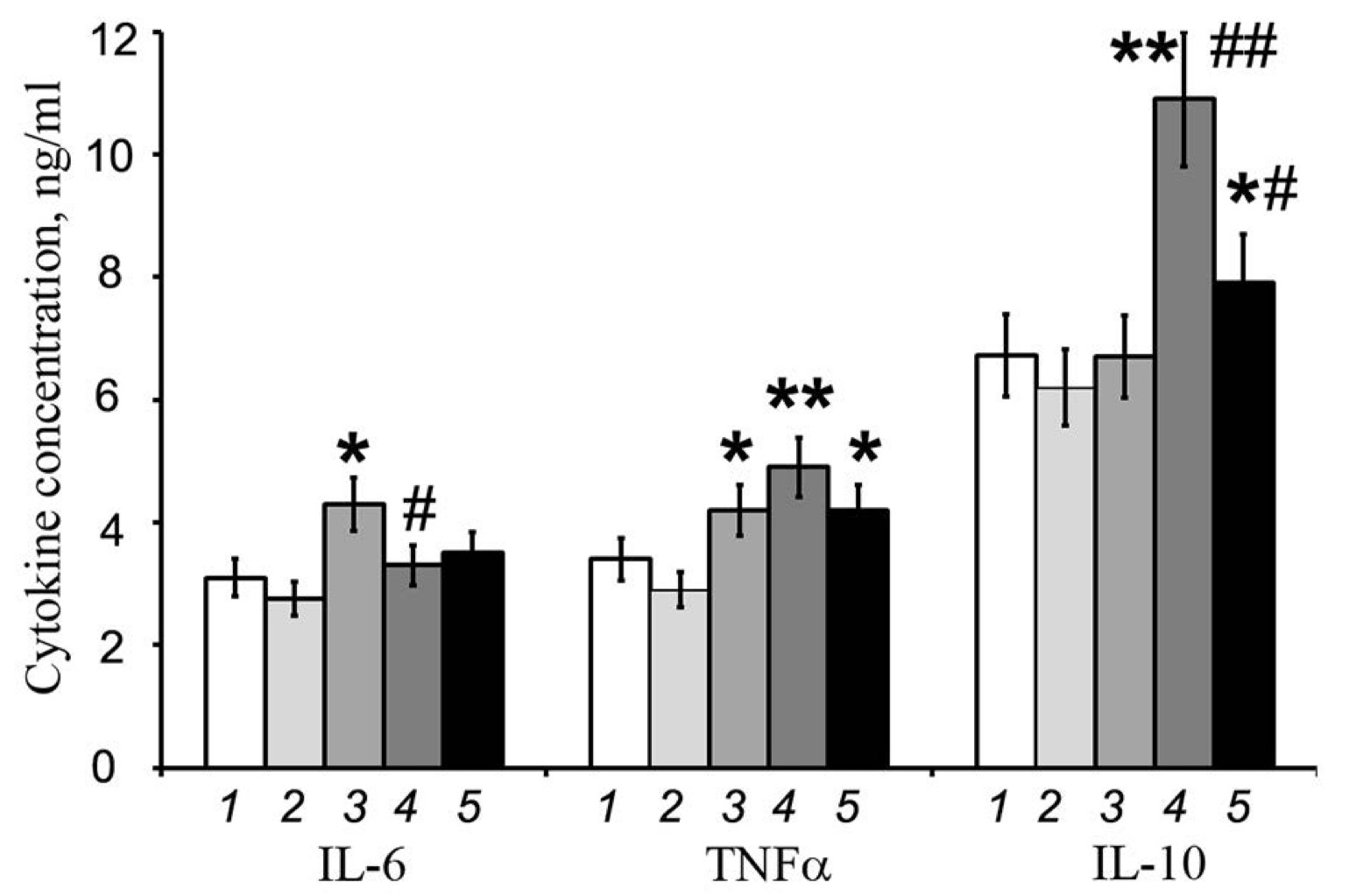
Fig. 4: Effect of streptozotocin and peroxiredoxin 6 on the production of the proinflammatory cytokines IL-6 and TNF-α and the anti-inflammatory cytokine IL-10 by RIN-m5F cells. The results of the ELISA analysis are presented. 1 - control, 2 - PRDX6, 3 - STZ, 4 - STZ-induced PRDX6 simultaneously, 5 - PRDX6 one hour before STZ. The values are the means of 3 independent experiments ± SEMs. *- significant difference from the control group (p<0.05), **- significant difference from the control group (p<0.01), # - significant difference from the STZ group (p<0.05), ## - significant difference from the STZ group (p <0.01)
Effects of PRDX6 and STZ on NF-κB pathway activation and heat shock protein production in RIN-m5F β-cells
When
culturing RIN-m5F cells in the presence of STZ, we detected a
decrease in the production of the protective inducible heat shock
protein HSP70 but not HSP90 (Fig. 2). Moreover, modification of NF-κB
signaling was observed: along with a strong increase in the total p65
protein content, its phosphorylation at serine 536 was significantly
reduced. The addition of peroxiredoxin 6 to β-cells resulted in the
activation of the NF-κB signaling pathway but not heat shock stress
proteins. By studying the mechanisms of the senotherapeutic action of
PRDX6, we found that the presence of this protein may reduce the
aggressive effect of STZ, activating the expression of the
stress-inducible heat shock proteins HSP90α and HSP70 but not the
constitutive form of HSP90β. In addition, preliminary administration
of the antioxidant protein prevented STZ-induced inhibition of p65
phosphorylation at serine 536.
Peroxiredoxin improves the viability of RIN-m5F β-cells in STZ-containing medium
In
addition, we assessed the number of live RIN-m5F β-cells after
culture with STZ and showed that, under these conditions, the number
of these cells was significantly reduced. However, when PRDX6 was
present in the culture medium of β-cells exposed to STZ, a
significant increase in the number of live RIN-m5F β-cells was
observed compared to that in the STZ group (Fig. 5).
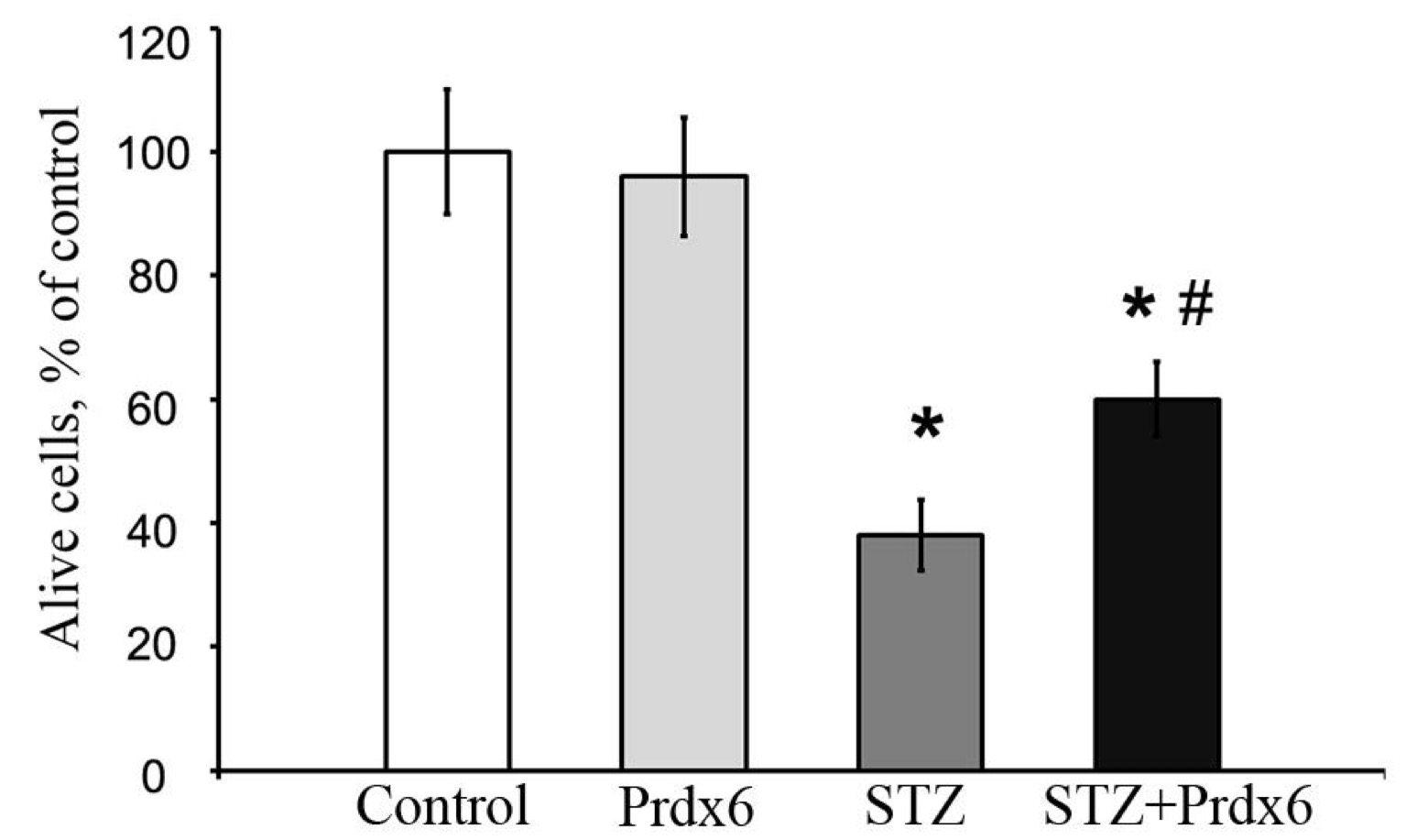
Fig. 5: Effect of streptozotocin and peroxiredoxin 6 on the viability of RIN-m5F cells after 24 hours of incubation. Data on the percentage of viable cells are presented. The number of live cells in the control sample was taken as 100%. Groups: 1 - control, 2 - PRDX6, 3 - STZ, 4 - simultaneous treatment with PRDX6 and STZ. The values are the means of 3 independent experiments ± SEMs. *- significant difference from the control group (p<0.05), # - significant difference from the STZ group (p<0.05), *# - significant difference from the STZ group (p<0.01).
Discussion
Due to the increase in the life expectancy and well-being of humans, metabolic processes associated with aging and age-associated diseases, such as obesity and type 2 diabetes, are currently a serious and socially significant concern. We previously established that PRDX6 exerts a protective effect against the development of type 1 diabetes mellitus as well as against ionizing radiation [22, 26]. However, increased insulin resistance and impaired insulin secretion are important characteristics of type 2 diabetes mellitus and may be associated with, among other factors, age-associated and stress-associated senescence of pancreatic insulin-producing cells. Targeting senescent cells may present many therapeutic opportunities, especially in the context of age-related diseases. This concept has stimulated the testing of senolytic and senomorphic drugs as a new therapeutic paradigm. The emergence of senescent β-cells leads to increased insulin resistance and impaired insulin secretion, which plays a central role in the pathophysiology of TD2 [27, 28].
Age-related changes in β-cell function and proliferation suggest the importance of senescent pancreatic β-cells in the pathogenesis of TD2. Cellular aging or senescence is a condition that develops as a result of replicative shortening of telomeres or the influence of genotoxic, oncogenic and oxidative stresses and is characterized by irreversible cell cycle arrest and cell dysfunctions. Senescent cells are characterized by a set of transcriptional, epigenetic, morphological and metabolic changes and produce a complex secretome (known as the senescence-associated secretory phenotype (SASP)), which causes senescence of surrounding cells (28, 29). Moreover, senescent cells may partially retain their main function, but they may be dysregulated in some aspects, accounting for major internal metabolic rearrangements occurring in these cells. Although many of the detrimental effects of aging can be explained by the SASP, direct evidence for the role of the aggressive phenotype is often lacking given the heterogeneity of the SASP and its context-dependent consequences. The nonspecific immune and heat shock stress protein (HSP) signaling pathways play important roles in cell senescence. These pathways are involved in the regulation of senescent cell survival and the development of the SASP and therefore may be targets for targeted senotherapy for age-associated diseases.
In our model of β-cell senescence, rat insulinoma RIN-m5F cells in the presence of the senescence-inducing agent STZ exhibited all the following signs of cell senescence: an increase in the level of oxidative stress, the percentage of SA-β-Gal-positive cells, the accumulation of the genotoxic stress marker phH2A and the universal inhibitor of the cellular p21 cycle, as well as an increase in the secretion of the proinflammatory cytokine IL-6 and impaired NF-κB phosphorylation. In addition, a decrease in the number of viable cells was observed when the cells were cultured with STZ. Therefore, STZ may be used for the development of cellular models of type 1 diabetes, which is well known, and also for the development of cellular models of type 2 diabetes, accounting for the abovementioned relationships between β-cell senescence and insulin resistance [27, 28].
In view of the role of oxidative stress in the induction of senescence, the aggressive effects of the SASP [29], and the overall vulnerability of β-cells to such harmful influences, the antioxidant enzyme peroxiredoxin 6, which has previously shown senomorphic effects, was used as a serotherapeutic agent in this model. Our data demonstrated that PRDX6, especially when added 1 hour before STZ, was able to prevent many of the pathological effects of STZ on RIN-m5F β-cells and decrease the number of senescent cells, indicating the potential protective role of PRDX6 in type 2 diabetes mellitus.
Identification of reliable markers of aging in vivo and models that allow for targeted manipulation of SASP factors inside senescent cells is an urgent task of modern cytogerontology. Therefore, when developing senotherapy for a specific age-associated disease, it is necessary to understand the characteristics of senescence of different types of cells and the possibility of reducing their aggressive secretome since the elimination of certain types of senescent cells can be fatal for living organisms [30-37]. Thus, β-cell senescence and the development of both type 1 and type 2 diabetes mellitus may have a common genetic basis. Indeed, diabetes mellitus has been reported to coexist with LMNA-mediated progeria and Werner syndrome [38, 39].
Targeting senescent cells to slow the aging process and reduce their level of dysfunction, known as “senotherapy,” is gaining momentum. The little-studied area associated with the clinical use of senotherapeutic drugs requires accelerated development. Further research into the molecular mechanisms of cellular senescence in diabetes and the clinical efficacy and safety of senolytic compounds should enable clinical application of this concept.
Acknowledgements
The authors thank AJE for help with language editing. The equipment used in the study (plate reader Infinite 200 (Tecan)) was obtained from the Collective Use Center of the Pushchino Scientific Center.
Funding
This work was supported by the Russian Science Foundation, grant No 23-24-00041.
Data availability
The data generated in the present study may be requested from the corresponding author
Authors' contributions
Concept
and management – E.G. Novoselova; planning and performing of
experiments – O.V. Glushkova, M.O. Khrenov, T.V. Novoselova, S.B.
Parfenyuk, S.M. Lunin. M.G. Sharapov, Discussion of the results –
C.M. Lunin, O.V. Glushkova, M.G. Sharapov. Text writing – E.G.
Novoselova. Text editing - S.M. Lunin, O.V. Glushkova, S.B.
Parfenyuk, M.O. Khrenov.
Disclosure Statement
The authors declare that they have no competing interests.
References
| 1 | King H, Aubert RE, Herman WH: Global burden of diabetes, 1995-2025: Prevalence, numerical estimates, and projections. Diabetes Care 1998;21:1414-1431.
https://doi.org/10.2337/diacare.21.9.1414 |
| 2 | IDF Diabetes Atlas, ed 9, Brussels, International Diabetes Federation, 2019.
|
| 3 | Pearson ER: Type 2 diabetes: a multifaceted disease. Diabetologia 2019;62:1107-1112.
https://doi.org/10.1007/s00125-019-4909-y |
| 4 | Donath MY, Shoelson SE: Type 2 diabetes as an inflammatory disease. Nat Rev Immunol 2011;11:98-107.
https://doi.org/10.1038/nri2925 |
| 5 | Christensen AA, Gannon M: The beta cell in type 2 diabetes. Curr Diab Rep 2019;19:81.
https://doi.org/10.1007/s11892-019-1196-4 |
| 6 | Rutter GA, Pullen TJ, Hodson DJ, Martinez-Sanchez A: Pancreatic β-cell identity, glucose sensing and the control of insulin secretion. Biochem J 2015;466:203-218.
https://doi.org/10.1042/BJ20141384 |
| 7 | Desai S, Deshmukh A, Mapping of type 1 diabetes mellitus. Curr Diabetes Rev 2020;16:438-441.
https://doi.org/10.2174/1573399815666191004112647 |
| 8 | Ilonen J, Lempainen J, Veijola R: The heterogeneous pathogenesis of type 1 diabetes mellitus. Nat Rev Endocrinol 2019;15:635-650.
https://doi.org/10.1038/s41574-019-0254-y |
| 9 | Yahaya TO, Salisu TF: A review of type 2 diabetes mellitus predisposing genes. Curr Diabetes Rev 2019;16:52-61.
https://doi.org/10.2174/1573399815666181204145806 |
| 10 | Lisco G, Giagulli VA, De Pergola G, Guastamacchia E, Jirillo E, Triggiani V: Pancreatic macrophages and their diabetogenic effects: highlight on several metabolic scenarios and dietary approach. Endocr Metab Immune Disord Drug Targets 2023;23:304-315.
https://doi.org/10.2174/1871530322666220510123913 |
| 11 | Malaisse WJ, Malaisse-Lagae, F, Sener A, Pipeleers DG: Determinants of the selective toxicity of alloxan to the pancreatic B cell. Proc Natl Acad Sci. USA 1982;79:927-930.
https://doi.org/10.1073/pnas.79.3.927 |
| 12 | Miranda-Díaz AG, Pazarín-Villaseñor L, Yanowsky-Escatell FG, Andrade-Sierra J: Oxidative stress in diabetic nephropathy with early chronic kidney disease. J Diabetes Res 2016;6:7047238.
https://doi.org/10.1155/2016/7047238 |
| 13 | Maiese K: New insights for oxidative stress and diabetes mellitus. Oxid Med Cell Longev 2015;2015:875961.
https://doi.org/10.1155/2015/875961 |
| 14 | Savastio S, Cadario F, Genoni G, Bellomo G, Bagnati M, Secco G, Picchi R, Giglione E, Bona G: Vitamin D deficiency and glycemic status in children and adolescents with type 1 diabetes mellitus. PLoS One 2016;11:e0162554.
https://doi.org/10.1371/journal.pone.0162554 |
| 15 | Ceriello A: New insights on oxidative stress and diabetic complications may lead to a "causal" antioxidant therapy. Diabetes Care 2003;26:1589-1596.
https://doi.org/10.2337/diacare.26.5.1589 |
| 16 | Boudina S, Abel ED: Diabetic cardiomyopathy revisited. Circulation 2007;115:3213-3223.
https://doi.org/10.1161/CIRCULATIONAHA.106.679597 |
| 17 | Navas LE, Carnero A: NAD+ metabolism, stemness, the immune response, and cancer. Signal Transduct Target Ther 2021;6:2.
https://doi.org/10.1038/s41392-020-00354-w |
| 18 | Szkudelski T: The mechanism of alloxan and streptozotocin action in B cells of the rat pancreas. Physiol Res 2001;50:537-546.
https://doi.org/10.33549/physiolres.930111 |
| 19 | Sharapov MG, Glushkova OV, Parfenyuk SB, Gudkov SV, Lunin SM, Novoselova EG,: The role of TLR4/NF-κB signaling in the radioprotective effects of exogenous Prdx6. Arch Biochem Biophys 2021;702:108830.
https://doi.org/10.1016/j.abb.2021.108830 |
| 20 | Sharapov MG, Goncharov RG, Parfenyuk SB, Glushkova OV, Novoselov VI: The role of phospholipase activity of peroxiredoxin 6 in its transmembrane transport and protective properties. Int J Mol Sci 2022;23:15265.
https://doi.org/10.3390/ijms232315265 |
| 21 | Garay-Malpartida HM, Mourão RF, Mantovani M, Santos IA, Sogayar MC, Goldberg AC: Toll-like receptor 4 (TLR4) expression in human and murine pancreatic beta-cells affects cell viability and insulin homeostasis. BMC Immunol 2011;12:18.
https://doi.org/10.1186/1471-2172-12-18 |
| 22 | Novoselova EG, Sharapov MG, Lunin SM, Parfenyuk, SB, Khrenov MO, Mubarakshina EK, Kuzekova AA, Novoselova TV, Goncharov RG, Glushkova OV: Peroxiredoxin 6 applied after exposure attenuates damaging effects of X-ray radiation in 3T3 mouse fibroblasts. Antioxidants 2021;10:1951.
https://doi.org/10.3390/antiox10121951 |
| 23 | Sharapov, MG, Novoselov VI, Fesenko EE, Bruskov VI, Gudkov SV: The role of peroxiredoxin 6 in neutralization of X-ray mediated oxidative stress: effects on gene expression, preservation of radiosensitive tissues and postradiation survival of animals. Free Radical Research 2017;51:148-166.
https://doi.org/10.1080/10715762.2017.1289377 |
| 24 | Sharapov MG, Novoselov VI, Ravin VK: Cloning, expression and comparative analysis of peroxiredoxin 6 from different species. Mol Biol 2009;43:505-511.
https://doi.org/10.1134/S0026893309030145 |
| 25 | Itahana K, Campisi J, Dimri GP: Methods to detect biomarkers of cellular senescence: the senescence-associated beta-galactosidase assay. Methods Mol Biol 2007;371:21-31.
https://doi.org/10.1007/978-1-59745-361-5_3 |
| 26 | Novoselova EG, Glushkova OV, Lunin SM, Khrenov, MO, Parfenyuk SB, Novoselova TV, Sharapov MG, Gordeeva AE, Novoselov VI, Fesenko EE: Thymulin and peroxiredoxin 6 have protective effects against streptozotocin-induced type 1 diabetes in mice. Int J Immunopathol Pharmacol 2021;35:20587384211005645.
https://doi.org/10.1177/20587384211005645 |
| 27 | Novoselova EG, Lunin SM, Khrenov MO, Glushkova OV, Novoselova TV, Parfenyuk SB: The possible role of Β-cell senescence in the development of type 2 diabetes mellitus. Cell Physiol Biochem 2023;57:34-48.
https://doi.org/10.33594/000000606 |
| 28 | Xu M, Pirtskhalava T, Farr JN, Weigand BM, Palmer AK, Weivoda MM, Inman CL: Senolytics improve physical function and increase lifespan in old age. Nat Med 201;24:1246-1256.
https://doi.org/10.1038/s41591-018-0092-9 |
| 29 | Coppé JP, Patil CK, Rodier F, Sun Y, Muñoz DP, Goldstein J, Nelson PS, Desprez PY, Campisi J: Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol 2008;6:2853-2868.
https://doi.org/10.1371/journal.pbio.0060301 |
| 30 | Xu M, Tchkonia T, Ding H, Ogrodnik M, Lubbers ER, Pirtskhalava T, White TA, Johnson KO, Stout MB, Mezera V: JAK inhibition alleviates the cellular senescence associated secretory phenotype and frailty in old age. Proc Natl Acad Sci U S A 2015;112:6301-6310.
https://doi.org/10.1073/pnas.1515386112 |
| 31 | Murakami T, Inagaki N, Kondoh H: Cellular senescence in diabetes mellitus: distinct senotherapeutic strategies for adipose tissue and pancreatic b cells. Front Endocrinol 2022;13:869414.
https://doi.org/10.3389/fendo.2022.869414 |
| 32 | Rhodes CJ: Type 2 diabetes-a matter of beta-cell life and death? Science 2005;307:380-384.
https://doi.org/10.1126/science.1104345 |
| 33 | Childs BG, Durik M, Baker DJ, van Deursen JM: Cellular senescence in aging and age-related disease: from mechanisms to therapy. Nat Med 2015;21:1424-1435.
https://doi.org/10.1038/nm.4000 |
| 34 | Gorgoulis V, Adams PD, Alimonti A, Bennett DC, Bischof O, Bishop C, Campisi J, Collado M, Evangelou K, Ferbeyre G: Cellular senescence: defining a path forward. Cell 2019;179:813-827.
https://doi.org/10.1016/j.cell.2019.10.005 |
| 35 | Grosse L, Wagner N, Emelyanov A, Molina C, Lacas-Gervais S, Wagner KD, Bulavin DV: Defined p16(high) senescent cell types are indispensable for mouse healthspan. Cell Metab 2020;32:87-99.
https://doi.org/10.1016/j.cmet.2020.05.002 |
| 36 | Kang TW, Yevsa T, Woller N, Hoenicke L, Wuestefeld T, Dauch D, Hohmeyer A, Gereke M, Rudalska R, Potapova A: Senescence surveillance of pre-malignant hepatocytes limits liver cancer development. Nature 2011;479:547-551.
https://doi.org/10.1038/nature10599 |
| 37 | Midha A, Pan H, Abarca C, Andle J, Carapeto P, Bonner-Weir S, Aguayo-Mazzucato C: Unique human and mouse β-cell senescence-associated secretory phenotype (sasp) reveal conserved signaling pathways and heterogeneous factors. Diabetes 2021;70:1098-1116.
https://doi.org/10.2337/db20-0553 |
| 38 | He G, Yan Z, Sun L, Lv Y, Guo W, Gang X, Wang G: Diabetes mellitus coexisted with progeria: a case report of atypical Werner syndrome with novel LMNA mutations and literature review. Endocr J 2019;66:961-969.
https://doi.org/10.1507/endocrj.EJ19-0014 |
| 39 | Donadille B, D'Anella P, Auclair M, Uhrhammer N, Sorel M, Grigorescu R, Ouzounian S, Cambonie G, Boulot P, Laforêt P: Partial lipodystrophy with severe insulin resistance and adult progeria Werner syndrome. Orphanet J Rare Dis 2013;8:106.
https://doi.org/10.1186/1750-1172-8-106 |