Cardiotoxicity Induced by Intratracheal Instillation of Diesel Exhaust Particles in Mice, and the Protective Effects of Carnosol: Suppression of Inflammation and Oxidative and Nitrosative Stress via Modulation of NF-κb/MAPKs Signaling Pathways
Keywords
Abstract
Background/Aims:
Inhaled particulate air pollution is associated with cardiotoxicity with underlying mechanisms including oxidative stress and inflammation. Carnosol, commonly found in rosemary and sage, is known to possess a broad range of therapeutic properties such as antioxidant, anti-inflammatory and antiapoptotic. However, its cardioprotective effects on diesel exhaust particles (DEPs)-induced toxicity have not been studied yet. Hence, we evaluated the potential ameliorative effects of carnosol on DEPs-induced heart toxicity in mice, and the underlying mechanisms involved.Methods:
Mice were intratracheally instilled with DEPs (1 mg/kg) or saline, and 1 hour prior to instillation they were given intraperitoneally either carnosol (20 mg/kg) or saline. Twenty-four hours after the DEPs instillation, multiple parameters were evaluated in the heart by enzyme-linked immunosorbent assay, colorimetric assay, Comet assay and Western blot technique.Results:
Carnosol has significantly reduced the elevation in the plasma levels of lactate hydrogenase and brain natriuretic peptide induced by DEPs. Likewise, the augmented cardiac levels of proinflammatory cytokines, lipid peroxidation, and total nitric oxide in DEPs-treated groups were significantly normalized with the treatment of carnosol. Moreover, carnosol has markedly reduced the heart mitochondrial dysfunction, as well as DNA damage and apoptosis of mice treated with DEPs. Similarly, carnosol significantly reduced the elevated expressions of phosphorylated nuclear factor-кB (NF-кB) and mitogen-activated protein kinases (MAPKs) in the hearts. Furthermore, the treatment with carnosol has restored the decrease in the expression of sirtuin-1 in the hearts of mice exposed to DEPs.Conclusion:
Carnosol significantly attenuated DEP-induced cardiotoxicity in mice by suppressing inflammation, oxidative stress, DNA damage, and apoptosis, at least partly via mechanisms involving sirtuin-1 activation and the inhibition of NF-кB and MAPKs activation.Introduction
Air pollution is considered one of the most important risk factors for acute cardiovascular events and is associated with over 8.9 deaths annually, with ischemic heart disease and stroke accounting for half of the premature cardiovascular-related death [1-3]. Despite improvement in air quality in cities in the West, urban air pollution is still a pressing issue, particularly in developing countries where urbanization is at its peak [1, 2]. In addition to traffic vehicles, high-density residential and industrial areas are major emitters of anthropogenic gaseous, semi-volatile liquids, and particulate pollutants [2, 4]. These pollutants, especially particulate matter (PM) have been consistently associated with various detrimental cardiovascular effects, including cardiac arrest, arrhythmia and cerebrovascular disease [2]. PM is a concoction of particles from various sources and the smallest, yet the most volatile of all PMs is classified as PM0.1 (particles with a diameter less than 100 nm). This includes the byproduct of traffic-derived fossil fuel combustion, namely the diesel exhaust particles (DEPs), which is the most significant contributor to urban PM in developing countries and have been linked to various detrimental cardiac outcomes [5].
There are numerous mechanisms that mediate the adverse cardiovascular effects of particulate air pollution. However, previous studies have elucidated that oxidative stress and inflammation are the focal pathways involved [4, 6]. We have previously shown that acute pulmonary exposure to DEPs in mice is associated with cardiovascular dysfunctions [5, 7]. These effects are remarkably similar to those observed in both epidemiological and human controlled exposure studies and have shown comparable oxidative stress and inflammatory changes [8]. Moreover, the cardiovascular effects induced by inhaled air pollution often implicate the functions of other organs as well, hence, increasing both cardiovascular and all-cause mortality related to air pollution [1]. Therefore, it is essential to find therapeutic agents, including natural phytochemicals, with known antioxidant and anti-inflammatory properties to either halt or lessen the cardiotoxicity effects of particulate air pollution. A number of studies in the past, including ours, have attempted to use free radical scavengers and anti-inflammatory agents such omega-3 fatty acid, melatonin supplementations, as well as the phytochemicals catalpol and emodin to reduce the cardiovascular effects of air pollution, and have generated promising results [7, 9-11].
The antioxidant and anti-microbial properties of carnosol, a natural phenolic terpene profoundly found in rosemary (Rosemarinus officinalis) and sage (Salvia officinalis) have been established and widely applied in the food and cosmetics industries [12]. More specifically, several studies have shown that carnosol displays antioxidant, anti-inflammatory, antiapoptotic, antitumor and antithrombotic actions both in vitro and in vivo [13-16]. Therefore, given that a substantial body of evidence has shown that cardiotoxicity induced by DEPs is modulated by oxidative stress and inflammation, and carnosol is a potent antioxidant and anti-inflammatory agent, we thought that it would be enticing to assess the effectiveness of carnosol in attenuating cardiotoxicity induced by DEPs, which has not been reported yet. Consequently, here, we aim to evaluate the potential ameliorative effect of carnosol in preventing cardiotoxicity induced by DEPs and the mechanisms associated with it, by assessing various markers of oxidative stress, inflammation, mitochondrial dysfunction, DNA damage, and apoptosis. Furthermore, we wanted to investigate the involvement of other key mechanisms that regulate inflammatory responses by evaluating the expression of proteins involved in the mitogen-activated protein kinase (MAPK), nuclear factor-кB (NF-кB), and sirtuin-1 pathways.
Materials and Methods
Animals and DEPs
An
equal number of male and female BALB/c mice (Research Animal
Facility, College of Medicine and Health Sciences, UAE University, Al
Ain, United Arab Emirates) aged six to eight weeks weighing 25 – 30
g were housed in 12-hour light-dark cycle and temperature-controlled
(22 ± 1 °C) rooms. They had free access to commercially available
additive-free laboratory chow, which contained 24% of crude protein,
2% of crude fat, and 8% of crude fiber (National Feed and Flour and
Marketing Co., Abu Dhabi, United Arab Emirates) and filtered water.
The
DEPs were acquired from the National Institute of Standards and
Technology (Gaithersburg, MD, USA) and suspended in sterile saline
(NaCl 0.9%) with Tween 80 (0.01%). The DEP suspensions were sonicated
for 15 minutes and vigorously vortex upon suspension and diluted
prior to intratracheal (i.t.) instillation. These particles have been
analyzed by transmission electron microscopy and the presence of
small carbonaceous particle aggregates (<100 nm) was detected [5].
The majority of the aggregates were 1 µm in diameter or less and the
analysis of the geometric mean aerodynamic diameter of 215 nm was
reported previously from the same DEP material [11].
Mice Treatments and Carnosol
The
pulmonary exposure of DEPs to the mice was performed via
intratracheal (i.t.) instillation. Immediately before the
instillation mice were anesthetized with 5% isoflurane using the
Surgivet® model 100 vaporizer from ICU Medical, Inc. (San Clemente,
CA, USA) and placed supine on an angled board. A 24 G catheter was
used to instill 100 µl of either DEP (1 mg/kg) or saline
intratracheally and followed by 100 µl of air [7,
16, 17].
Twenty mg/kg of carnosol purchased from Santa Cruz Biotechnology,
Inc. (Dallas, TX, USA) was given intraperitoneally to the mice 1 hour
before the DEPs instillation [7,
11, 15, 16, 18].
The mice were randomly segregated into four groups and were treated
as follows:
- Saline – Treated with saline (10 mL/kg) via intraperitoneal (i.p.) injection 1 hour before the i.t. instillation of saline (100 µl).
- DEP – Treated with saline (10 mL/kg) via i.p. injection 1 hour before the i.t. instillation of DEP (1 mg/kg).
- Carnosol + Saline – Treated with carnosol (20 mg/kg) via i.p. injection 1 hour before the i.t. instillation of saline (100 µl).
- Carnosol + DEP – Treated with carnosol (20 mg/kg) via i.p. injection 1 hour before the i.t. instillation of DEP (1 mg/kg).
Sample Homogenization and Protein Estimation
The
frozen heart tissues were thawed and homogenized with 2.0 mm zirconia
beads from BioSpec (Bartlesville, OK, USA) in 2 mL microcentrifuge
tubes along with potassium chloride buffer supplemented with protease
and phosphatase inhibitors. The samples were homogenized for three
times of three cycles of 10 seconds at 6500x g
using
Precellys® homogenizer from Bertin Instruments (Bretonneux, France)
and then centrifuged at 14, 000x g
for 20 minutes. Subsequently, the supernatants were aliquoted into 4
tubes and kept at -80 °C pending further analysis.
The
protein quantification analysis was done as per the protocol that
came along with the Pierce™ Protein Assay Kit from Thermo
Scientific (Rockford, IL, USA).
Measurement
of Lactate Dehydrogenase (LDH) and Brain Natriuretic Peptide (BNP) in
Plasma
The
quantification of LDH in plasma was done as per the protocol
previously described [21].
In a 96-well plate, 50 µl of plasma and 50 µl of assay reagent
containing 4 mM iodonitrotetrazolium chloride in 0.2 M Tris-HCl (pH
8.2), 6.4 mM beta-nicotinamide adenine dinucleotide sodium salt and
320 mM lithium lactate in 0.2 M Tris-HCl (pH 8.2), along with 0.5 µl
of 150 mM 1-methoxyphenazine methosulfate in Tris-HCL (pH 8.2). Next,
the plate was briefly mixed on an orbital shaker for 15 seconds and
incubated in a dark for 60 minutes at room temperature. Subsequently,
the reaction was stopped by adding 50 µl of 1M acetic acid and the
absorbance was read and recorded at 490 nm. The plasma level of BNP,
a marker of cardiac dysfunction, was measured using an ELISA kit from
MyBioSource (San Diego, CA, USA).
Assessment of Proinflammatory Cytokines in Heart Homogenates
The concentrations of tumor necrotic factor α (TNFα), interleukin
(IL)-6, and IL-1β in the heart homogenates were assessed using ELISA
kits purchased from R&D Systems (Minneapolis, MN, USA) [22].
Measurement of Lipid Peroxidation and total nitric oxide (NO) in Heart Homogenates
The end product of lipid peroxidation, thiobarbituric acid reactive
substances (TBARS) was measured spectrophotometrically using
malondialdehyde from Sigma Aldrich Co. (St. Louis, MO, USA) as the
standard. Whereas, the NO activity was measured with a total NO assay
kit (R&D systems, Minneapolis, MN, United States), which was used
to quantify the more stable NO metabolites, NO2−
and NO3−.
Mitochondrial Extraction and Assessment of Mitochondrial Complexes in Heart Homogenates
The
isolation of mitochondria from the heart was done as previously
described via differential centrifugation, where the hearts were
chopped and homogenized in mitochondrial isolation buffer (MIB)
containing 0.32 M sucrose, 1 mM EDTA, 10 mM tris base [23,
24].
Subsequently, the homogenates were spun at 1000 rpm at 4° C for 10
minutes to remove cellular debris. Next, the supernatants were
centrifuged at 15, 000 rpm at 4° C for 15 minutes and pellets
containing mitochondria were resuspended in MIB, aliquoted, and kept
at -80° C for further use.
The
mitochondrial complex I, complex II & III, and complex IV
activities in the hearts of mice were assessed based on a protocol
previously published [24,
25].
For the analysis of rotenone-sensitive complex I, 25 µg of extracted
mitochondria was added to a 96-well plate along with distilled water,
10 mM potassium cyanide (KCN), 0.5 M potassium phosphate buffer (PPB,
pH 7.5), 10 mM NADH and 50 mg/mL BSA. In order to monitor the
specificity of this assay, parallelly in a separate plate, the same
reagents and samples were added as well as 1 mM rotenone. The
reaction was started by adding 5 mM of ubiquinone and the reduction
in absorbance at 340 nm was monitored for 10 minutes. The enzymatic
activity of complex I was calculated according to following the
equation: enzyme activity (nmol min-1mg-1)
= (Δ Absorbance/minx1000)/[(extinction coefficient x volume of
sample used in mL) x (sample protein concentration in mg mL-1)]
(extinction coefficient of NADH = 6.2 mM-1cm-1).
The
complex II & III activity was measured by adding distilled water,
0.5 M PPB (pH7.5), 10 mM KCN, 400 mM succinate solution, and 25 µg
extracted mitochondria into a 96-well plate. Next, the plate was
incubated on an orbital shaker for 10 min at 37° C. Reaction was
initiated with the addition of 1 mM oxidized cytochrome C and the
increase of absorbance was followed for 5 min at 550 nm. The
enzymatic specificity of the assay was determined by adding 1 M
malonate or 1 mg/mL antimycin A in another plate with the same
reagents and samples. The activity of complex II & III was
calculated using the same equation used for complex I (extinction
coefficient of reduced cytochrome C = 18.5 mM-1mg-1).
The
activity of complex IV was measured by adding distilled water, 0.1M
PPB (pH 7) and 1 mM reduced cytochrome C in a 96-well plate, followed
by the measurement of baseline activity at 550 nm. The reaction was
initiated by the addition of 25 µg of extracted mitochondria and the
reduction of absorbance at 550 nm was monitored for 10 min. In
parallel, 10 nM KCN was added in a separate plate containing the same
reagents and samples to evaluate the specificity of the assay. The
enzymatic activity of complex IV was calculated using the same
formula used for complex II & III.
DNA Damage in Heart
In
a separate set of mice, the hearts of mice from all groups were
excised immediately after sacrifice and were subjected to DNA damage
analysis as per the standard Comet assay protocol [26-28].
The DNA damage was assessed by measuring the length of the DNA
migration, including the nucleus diameter and the migrated DNA using
image analysis Axiovision 3.1 software by Carl Zeiss (White Plains,
NY, USA).
Measurement of Cleaved Caspase-3 in Heart Homogenates
The
level of cleaved caspase-3, in heart homogenates was measured using
an ELISA kit purchased from R&D Systems (Minneapolis, MN, USA)
Western
Blot Analysis of Phosphorylated (Phospho)-NF-κB and MAPKs in Heart
Homogenates
The
protein expressions of phospho-NF-кB and MAPKs including
phosphorylated c-Jun N-terminal kinase (phospho-JNK), phosphorylated
extracellular signal-regulated kinase (phospho-ERK) and
phosphorylated P38 (phospho-P38) were determined using the Western
blot technique. Thirty micrograms of protein from the heart
homogenates were separated in 10% sodium dodecyl sulfate
polyacrylamide gel via electrophoresis and then transferred onto
polyvinylidene difluoride membranes using the Trans-Blot Turbo
Transfer System from Bio-Rad (Hercules, CA, USA). Next, the blots
were incubated with 5% bovine serum albumin at room temperature for
one hour, before being probed with 1:1000 dilution of mouse
monoclonal phospho-NF-кB, p-JNK, p-ERK, and p-P38 antibodies from
Santa Cruz Biotechnology (Dallas, TX, USA) overnight at 4° C. The
next day, a 1:10, 000 dilution of rabbit anti-mouse IgG horseradish
peroxidase-conjugated secondary antibody from Abcam (Boston, MA, USA)
was used to incubate the blots for 2 hours at RT and the blots were
developed using the SuperSignal™ West Pico PLUS chemiluminescent
substrate from Thermo Scientific (Rockford, IL, USA). The measurement
of the protein band density was done using the image processing
program ImageJ (Bethesda, MD, USA). Blots were then re-probed with
1:10, 000 dilution of glyceraldehyde 3-phosphate dehydrogenase
(GAPDH) antibody from Abcam (Boston, MA, USA) and the protein band
density was used as the control.
Assessment
of the Concentration of Sirtuin-1 in Heart Homogenate
The
concentration of sirtuin-1 in heart homogenates was measured using an
ELISA kit purchased from MyBioSource (San Diego, CA, USA) as per the
protocol that came with the kit.
Statistics
The
statistical analyses were performed using GraphPad Prism Software
(GraphPad Software Inc., La Jolla, CA). Data are presented as mean ±
standard error of the mean, and statistical significance was
determined using one-way analysis of variance (ANOVA), followed by
the Holm–Sidak’s multiple comparisons test; p < 0.05 was
considered significant.
Results
LDH and BNP in Plasma
Fig. 1 shows that when compared with the saline group, treatment with DEPs, induced a significant increase in the levels of LDH (p<0.0001) and BNP (p<0.0001) in plasma. However, when carnosol was given an hour prior to DEPs instillation, the levels of LDH and BNP showed significant reduction (p<0.0001) compared with the group given only DEP.

Fig. 1: Activity of lactate dehydrogenase (LDH, A) and concentration brain natriuretic peptide (BNP, B) in plasma of mice, 24 h after intratracheal instillation of saline or diesel exhaust particles (DEPs, 1 mg/kg) with or without carnosol (20 mg/kg) pretreatment. Data are mean ± SEM (n=8). Statistical analysis by one-way analysis of variance followed by Holm-Sidak’s test.
TNFα, IL-6 and IL-1β in Heart Homogenates
Mice
i.t. instilled with DEP displayed a significant elevation in the
heart concentrations of inflammatory cytokines TNFα (p<0.0001),
IL-6 (p<0.0001)
and IL-1β (p<0.0001)
compared with the saline-treated mice. This elevation nevertheless
was significantly normalized (p<0.001,
p<0.0001
and p<0.0001,
respectively) with the pretreatment of carnosol (Fig. 2).
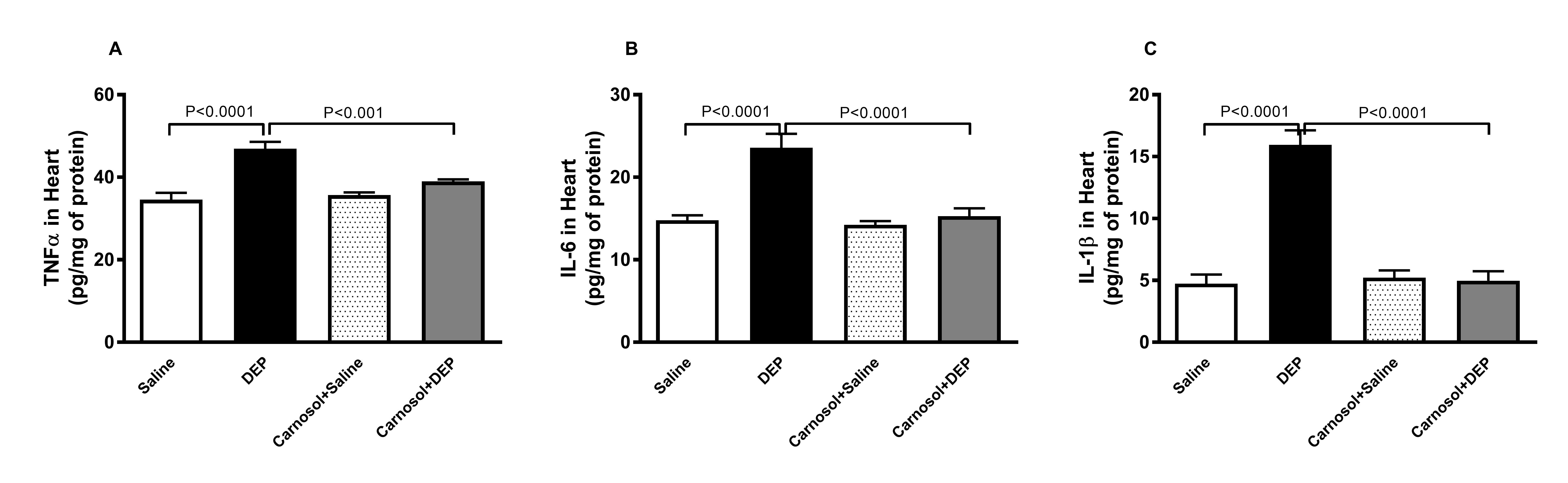
Fig. 2: Tumor necrosis factor α (TNFα, A), interleukin (IL)-6 (B) and IL-1β (C) concentrations in heart homogenates of mice, 24 hours after intratracheal instillation of saline or diesel exhaust particles (DEPs, 1 mg/kg) with or without carnosol (20 mg/kg) pretreatment. Data are mean ± SEM (n=8). Statistical analysis by one-way analysis of variance followed by Holm-Sidak’s test.
TBARS and NO in Heart Homogenates
Fig. 3 depicts that the changes in the levels of markers of oxidative
stress in heart homogenates in all four groups. The level of TBARS,
the byproduct of lipid peroxidation was significantly increased in
mice instilled with DEPs compared with the saline group (p<0.0001)
and this effect was significantly reduced (p<0.0001)
with the pretreatment with carnosol. Likewise, a significant increase
in NO activity (p<0.0001)
was recorded in DEPs-treated mice, in comparison with what is
observed in the saline group. This effect was significantly prevented
(p<0.0001)
when carnosol was given an hour before DEPs.
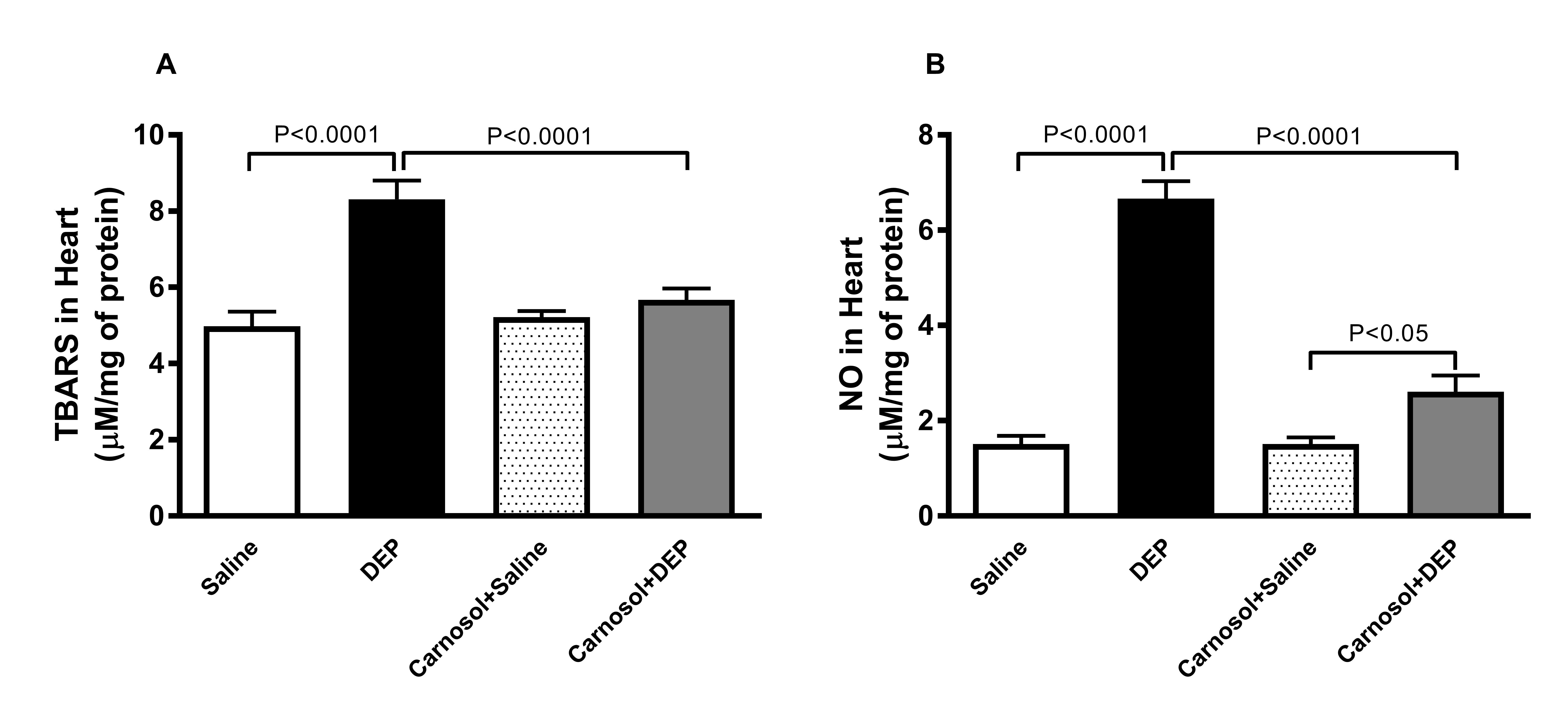
Fig. 3: Thiobarbituric acid reactive substances (TBARS, A) and total nitric oxide (NO, B) concentrations in heart homogenates of mice, 24 hours after intratracheal instillation of saline or diesel exhaust particles (DEPs, 1 mg/kg) with or without carnosol (20 mg/kg) pretreatment. Data are mean ± SEM (n=8). Statistical analysis by one-way analysis of variance followed by Holm-Sidak’s test.
Mitochondrial
Complex I, Complex II & III and Complex IV in Heart Homogenate
As
illustrated in Fig. 4, the activity of mitochondrial complex I,
complex II & III, and complex IV in the heart of mice with
pulmonary exposure to DEPs showed significant elevation (p<0.0001,
p<0.0001,
p<0.0001,
respectively) compared with the saline-treated mice. However, a
significant preventative effect was observed with the pretreatment of
carnosol one hour before DEPs (p<0.0001,
p<0.0001,
and p<0.0001,
respectively).

Fig. 4: Mitochondrial complexes I (A), II & III (B) and IV (C) activities in heart homogenates of mice, 24 hours after intratracheal instillation of saline or diesel exhaust particles (DEPs, 1 mg/kg) with or without carnosol (20 mg/kg) pretreatment. Data are mean ± SEM (n=8). Statistical analysis by one-way analysis of variance followed by Holm-Sidak’s test.
The DNA Damage in Heart
Fig.
5 shows that mice instilled with DEPs showed a significantly greater
level of DNA damage in the heart (p<0.0001)
compared with mice instilled with saline. This effect, however, was
significantly avoided (p<0.0001)
with carnosol pretreatment one hour prior to DEPs instillation.

Fig. 5: DNA migration (mm) in heart tissue by Comet assay in mice, 24 hours after intratracheal instillation of saline or diesel exhaust particles (DEPs, 1 mg/kg) with or without carnosol (20 mg/kg) pretreatment. Data are mean ± SEM (n=8). Statistical analysis by one-way analysis of variance followed by Holm-Sidak’s test.
Cleaved Caspase-3 in Heart Homogenate
When
compared with the saline group, pulmonary exposure to DEPs induced a
significantly higher concentration of the apoptotic marker cleaved
caspase-3 in hearts (p<0.0001,
Fig. 6). However, the pretreatment of carnosol significantly
normalized the concentration of cleaved caspase-3 in DEPs-treated
mice (p<0.0001).
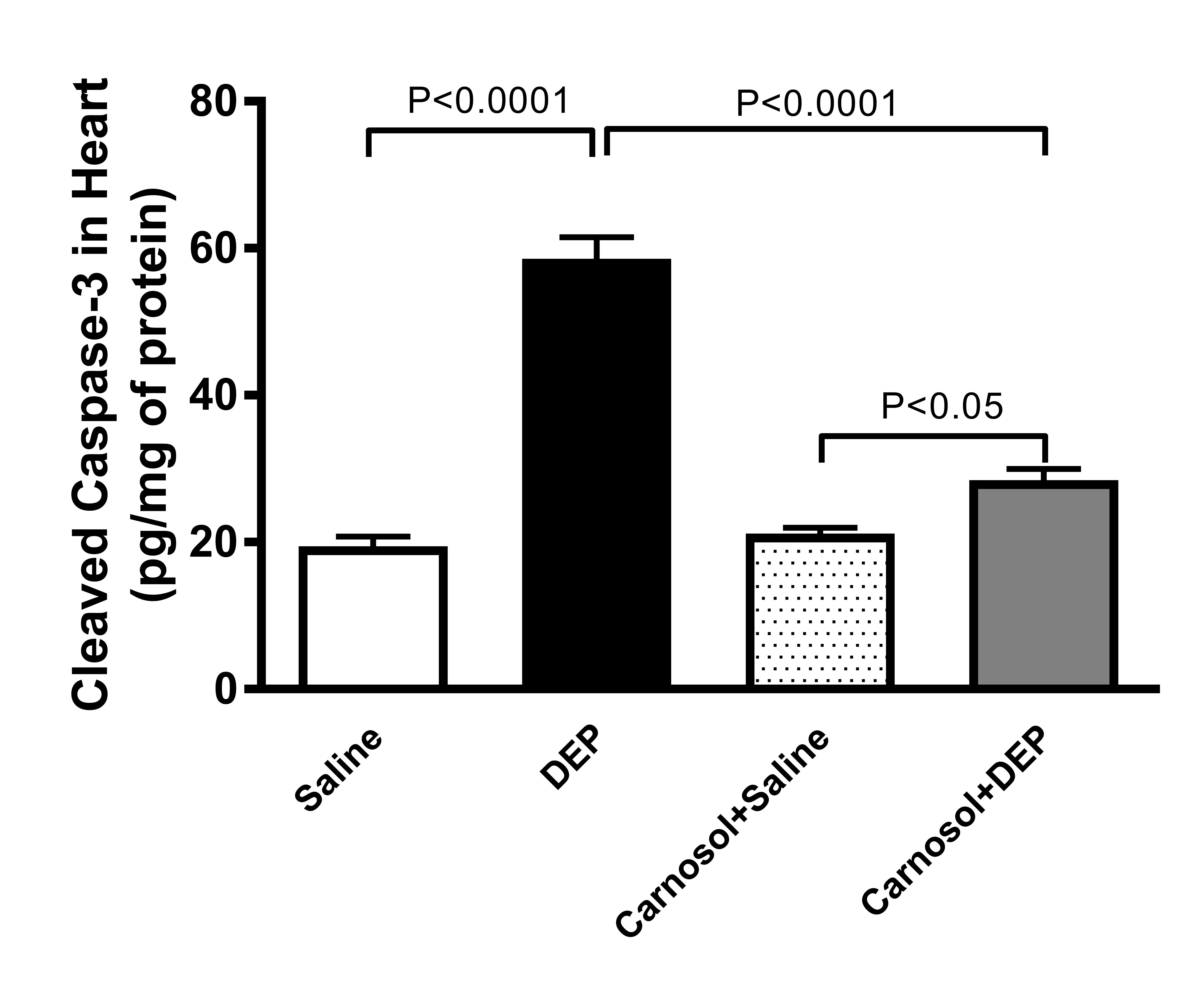
Fig. 6: Cleaved caspase-3 in heart homogenates of mice, 24 hours after intratracheal instillation of saline or diesel exhaust particles (DEPs, 1 mg/kg) with or without carnosol (20 mg/kg) pretreatment. Data are mean ± SEM (n=8). Statistical analysis by one-way analysis of variance followed by Holm-Sidak’s test.
Phospho-NF-κB in Heart Homogenate
Fig.
7 depicts the Western blot results related to the protein expression
of phospho-NF-кB in mice i.t. instilled with either DEPs or saline
with or without pretreatment with carnosol. DEPs exposure
significantly elevated the expression of phospho-NF-кB in the heart
of mice, compared with the saline group (p<0.0001).
This elevation nevertheless was significantly reversed when carnosol
pretreatment was introduced one hour before DEP instillation
(p<0.0001).
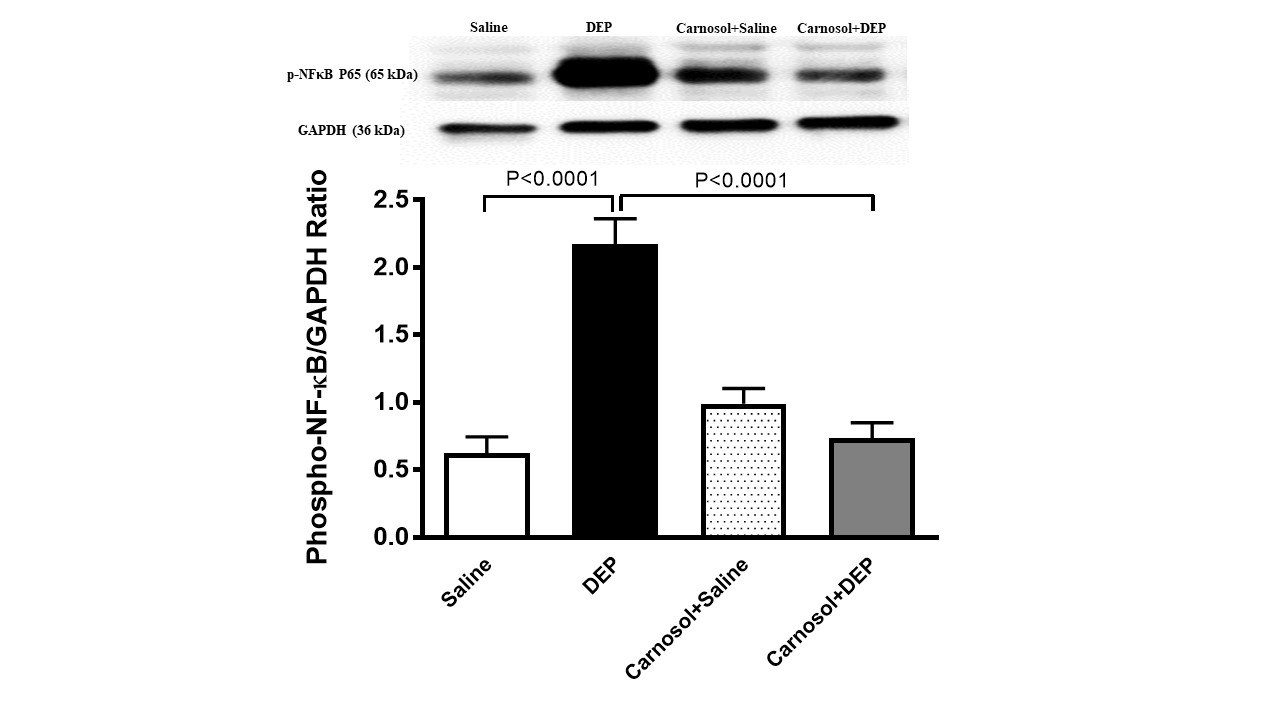
Fig. 7: Phosphorylated nuclear factor-кB (phosphor-NF-кB) expression in heart homogenates of mice, 24 hours after intratracheal instillation of saline or diesel exhaust particles (DEPs, 1 mg/kg) with or without carnosol (20 mg/kg) pretreatment. Data are mean ± SEM (n=8). Statistical analysis by one-way analysis of variance followed by Holm-Sidak’s test.
phospho-JNK, phospho-ERK, and phospho-38 in Heart Homogenate
The
Western blot analysis of the expression of markers of MAPKs is
illustrated in Fig. 8. Mice treated with DEPs showed significantly
higher expressions of phospho-JNK (p<0.0001),
phospho-ERK (p<0.0001),
and phospho-P38 (p<0.0001)
in hearts compared with mice treated with saline. The high
expressions of these markers, however, were significantly reversed by
treatment with carnosol (p<0.0001,
p<0.0001,
and p<0.0001,
respectively).
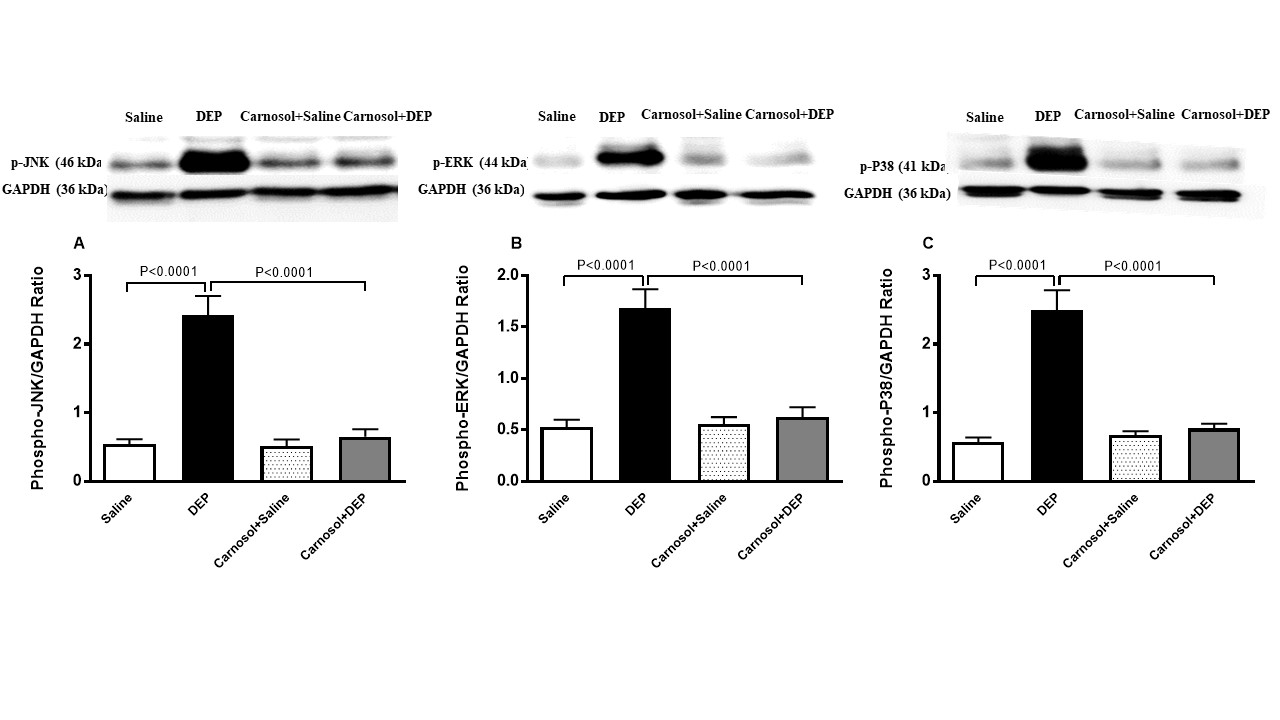
Fig. 8: Phosphorylated c-Jun N-terminal kinase (phospho-JNK, A), phosphorylated extracellular signal-regulated kinase (phospho-ERK, B) and phosphorylated P38 (phospho-P38, C) expression in heart homogenates of mice, 24 hours after intratracheal instillation of saline or diesel exhaust particles (DEPs, 1 mg/kg) with or without carnosol (20 mg/kg) pretreatment. Data are mean ± SEM (n=8). Statistical analysis by one-way analysis of variance followed by Holm-Sidak’s test.
Sirtuin in Heart Homogenate
Fig. 9 shows that the concentration of sirtuin-1 was markedly reduced in
DEPs-treated mice, compared with the saline treatment group
(p<0.0001).
An action that was significantly prevented in mice pretreated with
carnosol one hour prior to DEP instillation (p<0.001).
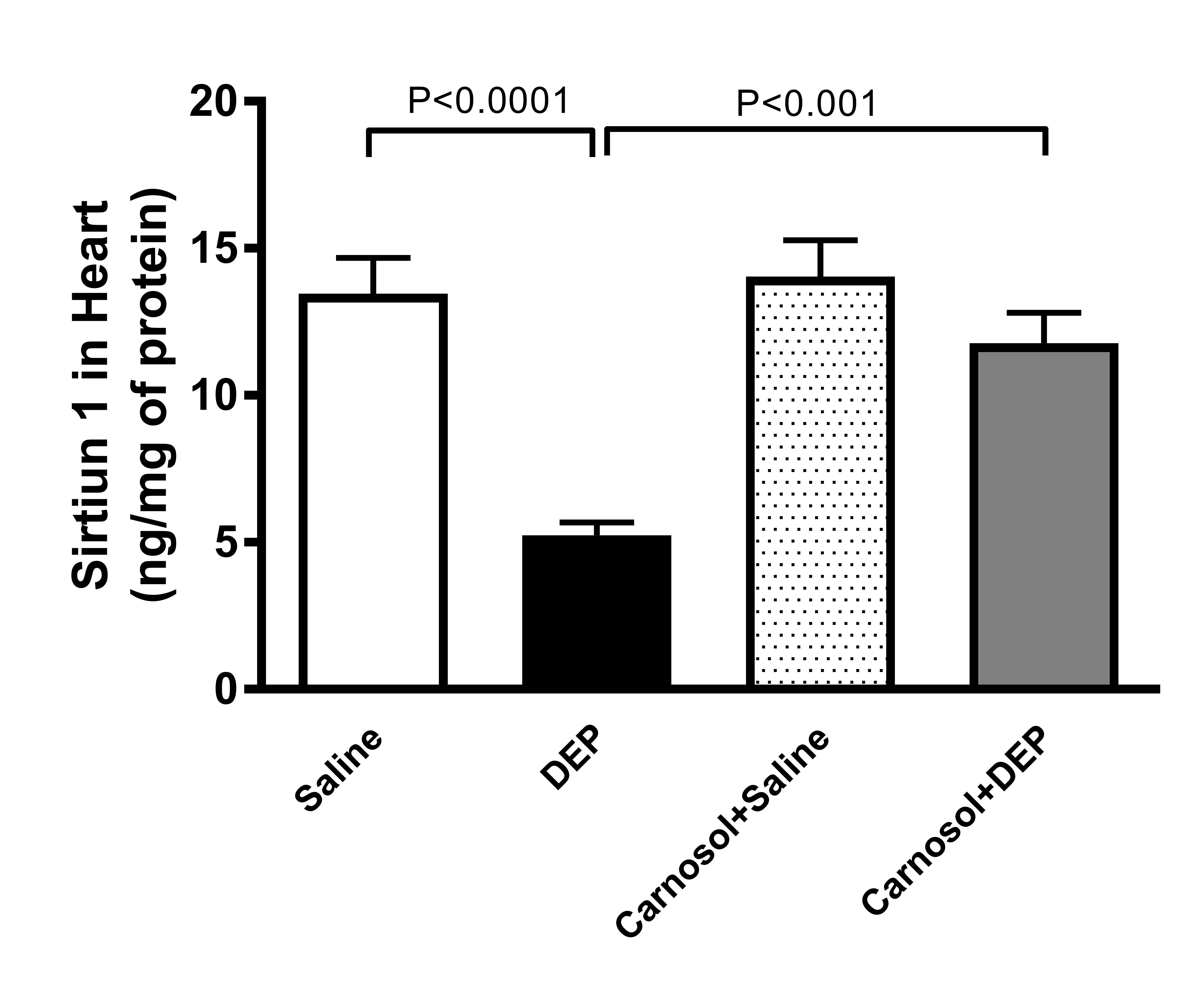
Fig. 9: Sirtuin-1 concentration in heart homogenates of mice, 24 hours after intratracheal instillation of saline or diesel exhaust particles (DEPs, 1 mg/kg) with or without carnosol (20 mg/kg) pretreatment. Data are mean ± SEM (n=8). Statistical analysis by one-way analysis of variance followed by Holm-Sidak’s test.
Discussion
In this study, we have demonstrated that pretreatment with carnosol has significantly attenuated the DEPs-induced cardiac inflammation, oxidative stress, mitochondrial dysfunction, DNA damage, and apoptosis. Moreover, carnosol has potently reversed the increase in the expressions of NF-кB and MAPKs and the decrease of sirtuin-1 in the heart.
There is now a substantial amount of evidence that links the exposure to air pollution with the detrimental effects on the cardiovascular system involving several interplaying mechanisms. Nevertheless, oxidative stress and inflammation have consistently been identified as the primary mechanisms involved in particulate air pollution-induced cardiotoxicity [7, 8]. Despite the wealth of knowledge on this topic, addressing particle air pollution from diesel exhaust remains challenging, particularly in developing countries where reliance on this efficient fossil fuel persists (up to 20% more fuel efficient than petrol) [29]. Legislative steps have been taken to reduce the global fossil fuel emissions, with many countries have introduced initiatives to phase out the reliance on fossil fuels, either by banning the sale of diesel and petrol vehicles, or by reducing the emissions, by replacing them with hybrid or electric cars [30]. However, these changes will take time and, in the meantime, it is best to consider other interventions including the use of phyto-antioxidants to reduce DEPs-induced oxidative stress and inflammation [1].
In the present study, we have pretreated mice intraperitoneally with 20 mg/kg carnosol, a phytochemical reported to have potent antioxidant and anti-inflammatory activities [31, 32]. The intraperitoneal administration is a widespread method of delivery in laboratory rodents and is recognized as a parenteral route of administration [33]. Thus the dosage and route of administration of carnosol applied here is comparable with other studies assessing its protective effects in animal models of human diseases [33]. It has been previously reported that carnosol is more efficient in neutralizing reactive oxygen species and has better DNA protecting ability than vitamin C and vitamin E [34]. Carnosol was given an hour before mice were intratracheally instilled with a bolus of DEPs suspension. This routine i.t. instillation method has been extensively applied in the past and could reproduce the adverse cardiovascular effects observed in particulate controlled exposure in human [5, 7, 8].
BNP is a sensitive marker for cardiac damage and LDH is a general marker of cytotoxicity [35-37]. The plasma concentration of BNP has been reported to be significantly elevated in an event of cardiac injury [35], a finding that is presently observed in mice instilled with DEPs, and confirmed by a previous report [24]. Both increases of BNP and LDH were averted by the pretreatment with carnosol. In conjunction with the latter observations, we wanted to appraise if the observed protective potential of carnosol is due to its ability to reduce oxidative stress, a key mediating pathway of DEPs-induced cardiotoxicity. Here, as previously reported [7], we have demonstrated that the level of the free radical scavenger NO was significantly augmented with DEPs instillation, an effect that was subsequently and significantly reduced with the pretreatment of carnosol. The increase in NO is perhaps the result of a compensatory process in which the heart produces more NO to defend itself from the oxidative stress, and the reduction of NO that came with carnosol pretreatment suggests that carnosol play an important role in eradicating DEPs-induced free radicals’ production [8, 38]. In order to produce a more robust evidence for the occurrence of oxidative stress, other alternate measures which reflect oxidative stress such as markers of lipid and DNA oxidation should be evaluated [8, 39]. In the present study, we have measured the concentration of TBARS, the byproduct of lipid peroxidation, and despite the increased in NO induced by DEPs, we found that the concentration of TBARS was significantly elevated in the heart of mice. This increase, however, was reversed with the pretreatment of carnosol, and concurred with a previous report that elucidated the ability of carnosol in preventing lipid peroxidation, by scavenging free radicals, as well as by directly inhibiting the lipid peroxidation process [31]. Moreover, in the present study, we have measured the DNA damage using Comet assay and detected an increase in DNA damage in DEPs-instilled mice, which is comparable to our precedent report [5]. Interestingly, a complete prevention in DNA damage was observed in the heart of mice pretreated with carnosol. These observations again suggest that carnosol might have cardioprotective ability by eliminating free radicals that could potentially oxidize both lipid and DNA. The oxidations of lipid and DNA have been linked to the increased buildup of oxidized lipids that would lead to inflammation, as well as oxidative DNA damage, that if not repaired, could give rise to several detrimental cellular responses [8, 38].
Although, the exact mechanisms by which inhaled particles caused adverse cardiovascular effects have not been fully elucidated yet, it is widely accepted that the interplay between oxidative stress and inflammation plays a mediating role in many pathological alterations including those that affect the heart in response to toxic agent exposure [8, 40]. It has been proposed that DEPs induce cardiovascular inflammatory response in three ways. First, by inducing inflammation in the lung, causing the secretion of cytokines into the circulatory system. Another hypothesis, suggests that DEPs directly induce inflammation by translocating across the air blood barrier into the circulatory system. The third proposal suggests that DEPs trigger sensory receptor activation causing alterations in the autonomic nervous system which subsequently lead to indirect functional changes to the cardiovascular system [8]. Regardless of the hypotheses, it is clear is that inflammation plays a pivotal role in DEP-induced cardiotoxicity. In the present study, we found that the DEPs-induced augmented levels of proinflammatory cytokines TNFα, IL-6 and IL-1β were significantly normalized with the pretreatment of carnosol. This suggest that carnosol is potentially effective in alleviating inflammation possibly via, if not all, at least one of the proposed pathways, and our data corroborate with previous reports where carnosol significantly reduced the concentrations of proinflammatory cytokines in murine models of acute lung injury and osteoporosis [18, 32].
The accumulation of DNA damage could lead to cascades of responses including apoptosis [38], an event characterized by the activation of caspase-3. Presently, DEPs significantly elevated the level of the activated form of caspase-3, the cleaved caspase-3, a finding that is consistent with our previous observation [7]. Remarkably the treatment with carnosol prevented this increase. The activation of cleaved caspase is prompted by both external and intracellular mechanisms [41]. The externally driven apoptosis involved the recruitment and the activation of caspase-3 by cytokines from the TNF superfamily including the TNFα, which we have shown earlier to be significantly normalized when carnosol is given an hour prior to DEPs exposure. While the internal cellular mechanism, activates the caspase-3 via the mitochondrial pathways [41]. It is possible that the reduction of caspase-3 is attributed to the efficiency of carnosol in averting its activation by preventing DNA damage and the expression of TNFα [42].
A number of epidemiological and observational studies have linked the etiology of particulate air pollution-induced cardiovascular toxicity to mitochondrial dysfunction [43]. These studies also suggest that mitochondria is the primary target of environmental toxicants which damage the morphology, functions and the mitochondrial DNA [43].. Moreover, mitochondrial dysfunction and defects have been linked to the increase of the generation of free radicals which could aggravate the oxidative stress and inflammation [24]. In this study, we have shown that the activities of mitochondrial complexes I, II&III and IV were significantly elevated when mice were given DEP, and amended when carnosol was given an hour prior. These suggest that carnosol could potentially protect mitochondria from particulate-induced cardiotoxicity and prevent the exacerbation of oxidative stress and inflammation associated with mitochondrial dysfunction. In fact, recent studies have reported that carnosol is effective in preventing mitochondrial dysfunction in models of nonalcoholic fatty liver disease and neurogenerative diseases [44, 45].
The NF-кB and MAPKs pathways are synergistically involved in the induction of pro-inflammatory cytokines and the progressions of many inflammatory diseases, including cardiovascular diseases, such as myocardial infarction, cardiac hypertrophy and atherosclerosis [7, 46-48]. Earlier studies have reported that carnosol is effective in mitigating inflammation associated with rheumatoid arthritis in mice, as well as in suppressing the inducible nitric oxide synthase involved in inflammation in mouse macrophages by downregulating the NF-кB and MAPK pathways [49, 50]. Here, we discovered that the significantly elevated expressions of phospho-NF-кB, and the markers of MAPKs (phospho-JNK, phospho-ERK and phospho-P38) induced by DEPs were significantly reduced with the pretreatment of carnosol. The reduction in phospho-NF-кB can be ascribed to the effectiveness of carnosol in inhibiting the activation of NF-кB by interfering with the signal-induced phosphorylation of IкB, which has been previously demonstrated in a transient transfection assay [50]. This subsequently, inhibits the production of proinflammatory cytokines. The carnosol-induced reduction of MAPKs markers suggests that carnosol could inhibit the phosphorylation of JNK, ERK and P38 MAPK, which attenuates the MAPKs signaling [51] and prevent the downstream inflammatory responses including the release of cytokines [52].
To further investigate the mechanism involved in the protective effects of carnosol in particulate induced cardiac inflammation, we have evaluated the concentration of sirtuin-1. Sirtuin-1 is an instrumental enzyme involved in the progression of cardiovascular disease, responsible in regulating the expression of inflammatory genes in the cells, as well as modulating lipid peroxidation, and the aging of endothelial progenitor cells [53, 54]. The activation of sirtuin-1 has been associated with improved oxidative metabolism and reduced inflammation, suggesting that sirtuin-1 has an inhibitory impact towards NF-кB signaling cascade [55]. Additionally, it has been previously reported that sirtuin-1 activation is linked to the decrease in phosphorylation of stress-induced JNK, ERK and P38, suggesting yet another inhibitory effect of sirtuin-1 on a pathway involved in inflammation [54, 56]. As a matter of fact, regulating the activity of sirtuin-1 has been applied pharmacologically under the notion that enhancing sirtuin-1 activity offers promising outcomes, which is evidence in various reports on the use of natural products with sirtuin-1 enhancing activity such as resveratrol, catalpol and quercetin [38, 53, 54]. Presently, we have found that carnosol has significantly abrogated the detrimental effect associated with the reduced level of sirtuin-1 found in the heart of mice treated with DEP, an observation that is concurred by previous claims where the therapeutic properties of carnosol via sirtuin-1 activation has been demonstrated on rats with liver fibrosis and early brain injury after subarachnoid hemorrhage [57, 58].
Conclusion
Taken together, our findings demonstrated that pretreatment with carnosol exhibited protective effects against DEPs-induced acute cardiotoxicity in mice by attenuating inflammation, oxidative stress, mitochondrial dysfunction, DNA damage and apoptosis. This was potentially achieved due to the ability of carnosol in activating sirtuin-1, which consequently suppressed the NF-кB and MAPK pathways. Nonetheless, further studies are warranted in order to substantiate this claim and to determine whether carnosol as a viable option for the clinical treatment of particulate-induced cardiotoxicity.
Acknowledgements
Funding
This
research was funded by Zayed Center for Health Sciences (grant number
12R166), College of Medicine and Health Sciences (grant numbers
12M112 and 12M167) and Summer Undergraduate Research Experience
(SURE) (grant number 2748) of the United Arab Emirates University.
Statement of Ethics
This
animal study protocol was approved by the Institutional Ethics
Committee of United Arab Emirates University (protocol code
ERA_2019)5876, approved on 9th April 2019).
Author Contributions
Conceptualization,
A.N.; Supervision, A.N., Methodology, N.E.Z., S.B., O.E. M.A. M.A.
and A.A.; Investigation, N.E.Z., S.B. and O.E.; Writing—original
draft preparation, N.E.Z.; Writing—review and editing, A.N.,
visualization, N.E.Z., S.B. and O.E, Funding Acquisition, A.N.
Disclosure Statement
The authors declare no conflicts of interest.
References
| 1 | Miller MR: The cardiovascular effects of air pollution: Prevention and reversal by pharmacological agents. Pharmacol Ther 2022;232:107996. DOI: 10.1016/j.pharmthera.2021.107996.
https://doi.org/10.1016/j.pharmthera.2021.107996 |
| 2 | Miller MR, Newby DE: Air pollution and cardiovascular disease: car sick. Cardiovasc Res 2020;116(2):279-94. DOI: 10.1093/cvr/cvz228.
https://doi.org/10.1093/cvr/cvz228 |
| 3 | Nemmar A, Hoylaerts MF, Nemery B: Effects of particulate air pollution on hemostasis. Clinics in occupational and environmental medicine 2006;5(4):865-81. DOI: 10.1016/j.coem.2006.07.007.
|
| 4 | Miller MR: Oxidative stress and the cardiovascular effects of air pollution. Free Radic Biol Med 2020;151:69-87. DOI: 10.1016/j.freeradbiomed.2020.01.004.
https://doi.org/10.1016/j.freeradbiomed.2020.01.004 |
| 5 | Nemmar A, Al-Salam S, Beegam S, Yuvaraju P, Ali BH: Thrombosis and systemic and cardiac oxidative stress and DNA damage induced by pulmonary exposure to diesel exhaust particles and the effect of nootkatone thereon. Am J Physiol Heart Circ Physiol 2018;314(5):H917-h27. DOI: 10.1152/ajpheart.00313.2017.
https://doi.org/10.1152/ajpheart.00313.2017 |
| 6 | Miller MR: The role of oxidative stress in the cardiovascular actions of particulate air pollution. Biochem Soc Trans 2014;42(4):1006-11. DOI: 10.1042/BST20140090.
https://doi.org/10.1042/BST20140090 |
| 7 | Nemmar A, Beegam S, Zaaba NE, Alblooshi S, Alseiari S, Ali BH: The Salutary Effects of Catalpol on Diesel Exhaust Particles-Induced Thrombogenic Changes and Cardiac Oxidative Stress, Inflammation and Apoptosis. Biomedicines 2022;10(1). DOI: 10.3390/biomedicines10010099.
https://doi.org/10.3390/biomedicines10010099 |
| 8 | Miller MR, Shaw CA, Langrish JP: From particles to patients: oxidative stress and the cardiovascular effects of air pollution. Future Cardiol 2012;8(4):577-602. DOI: 10.2217/fca.12.43.
https://doi.org/10.2217/fca.12.43 |
| 9 | Tong H, Rappold AG, Diaz-Sanchez D, Steck SE, Berntsen J, Cascio WE, Devlin RB, Samet JM: Omega-3 fatty acid supplementation appears to attenuate particulate air pollution-induced cardiac effects and lipid changes in healthy middle-aged adults. Environ Health Perspect 2012;120(7):952-7. DOI: 10.1289/ehp.1104472.
https://doi.org/10.1289/ehp.1104472 |
| 10 | Jiang J, Liang S, Zhang J, Du Z, Xu Q, Duan J, Sun Z: Melatonin ameliorates PM(2.5) -induced cardiac perivascular fibrosis through regulating mitochondrial redox homeostasis. J Pineal Res 2021;70(1):e12686. DOI: 10.1111/jpi.12686.
https://doi.org/10.1111/jpi.12686 |
| 11 | Nemmar A, Al Dhaheri R, Alamiri J, Al Hefeiti S, Al Saedi H, Beegam S, Yuvaraju P, Yasin J, Ali BH: Diesel Exhaust Particles Induce Impairment of Vascular and Cardiac Homeostasis in Mice: Ameliorative Effect of Emodin. Cell Physiol Biochem 2015;36(4):1517-26. DOI: 10.1159/000430315.
https://doi.org/10.1159/000430315 |
| 12 | Birtić S, Dussort P, Pierre FX, Bily AC, Roller M: Carnosic acid. Phytochemistry 2015;115:9-19. DOI: 10.1016/j.phytochem.2014.12.026.
https://doi.org/10.1016/j.phytochem.2014.12.026 |
| 13 | Sahu BD, Putcha UK, Kuncha M, Rachamalla SS, Sistla R: Carnosic acid promotes myocardial antioxidant response and prevents isoproterenol-induced myocardial oxidative stress and apoptosis in mice. Molecular and Cellular Biochemistry 2014;394(1):163-76. DOI: 10.1007/s11010-014-2092-5.
https://doi.org/10.1007/s11010-014-2092-5 |
| 14 | Baradaran Rahimi V, Momeni-Moghaddam MA, Chini MG, Saviano A, Maione F, Bifulco G, Rahmanian-Devin P, Jebalbarezy A, Askari VR: Carnosol Attenuates LPS-Induced Inflammation of Cardiomyoblasts by Inhibiting NF-κB: A Mechanistic in vitro and in Silico Study. Evid Based Complement Alternat Med 2022;2022:7969422. DOI: 10.1155/2022/7969422.
https://doi.org/10.1155/2022/7969422 |
| 15 | Rahnama M, Mahmoudi M, Zamani Taghizadeh Rabe S, Balali-Mood M, Karimi G, Tabasi N, Riahi-Zanjani B: Evaluation of anti-cancer and immunomodulatory effects of carnosol in a Balb/c WEHI-164 fibrosarcoma model. J Immunotoxicol 2015;12(3):231-8. DOI: 10.3109/1547691x.2014.934975.
https://doi.org/10.3109/1547691X.2014.934975 |
| 16 | Beegam S, Zaaba NE, Elzaki O, Alzaabi A, Alkaabi A, Alseiari K, Alshamsi N, Nemmar A: Palliative effects of carnosol on lung-deposited pollutant particles-induced thrombogenicity and vascular injury in mice. Pharmacology Research & Perspectives 2024;12(3):e1201. DOI: https://doi.org/10.1002/prp2.1201.
https://doi.org/10.1002/prp2.1201 |
| 17 | Nemmar A, Melghit K, Al-Salam S, Zia S, Dhanasekaran S, Attoub S, Al-Amri I, Ali BH: Acute respiratory and systemic toxicity of pulmonary exposure to rutile Fe-doped TiO2 nanorods. Toxicology 2011;279(1):167-75. DOI: https://doi.org/10.1016/j.tox.2010.10.007.
https://doi.org/10.1016/j.tox.2010.10.007 |
| 18 | Li Q, Liu L, Sun H, Cao K: Carnosic acid protects against lipopolysaccharide-induced acute lung injury in mice. Exp Ther Med 2019;18(5):3707-14. DOI: 10.3892/etm.2019.8042.
https://doi.org/10.3892/etm.2019.8042 |
| 19 | Ali BH, Ziada A, Husseni IA, Beegam S, Al-Ruqaishi B, Nemmar A: Effect of Acacia gum on blood pressure in rats with adenine-induced chronic renal failure. Phytomedicine 2011;18(13):1176-80. DOI: https://doi.org/10.1016/j.phymed.2011.03.005.
https://doi.org/10.1016/j.phymed.2011.03.005 |
| 20 | Benbarek H, Mouithys-Mickalad A, Deby-Dupont G, Deby C, Grülke S, Nemmar A, Lamy M, Serteyn D: High concentrations of histamine stimulate equine polymorphonuclear neutrophils to produce reactive oxygen species. Inflammation Research 1999;48(11):594-601. DOI: 10.1007/s000110050509.
https://doi.org/10.1007/s000110050509 |
| 21 | Kaja S, Payne AJ, Singh T, Ghuman JK, Sieck EG, Koulen P: An optimized lactate dehydrogenase release assay for screening of drug candidates in neuroscience. J Pharmacol Toxicol Methods 2015;73:1-6. DOI: 10.1016/j.vascn.2015.02.001.
https://doi.org/10.1016/j.vascn.2015.02.001 |
| 22 | Ali BH, Beegam S, Al-Lawati I, Waly MI, Al Za'abi M, Nemmar A: Comparative efficacy of three brands of gum acacia on adenine-induced chronic renal failure in rats. Physiol Res 2013;62(1):47-56. DOI: 10.33549/physiolres.932383.
https://doi.org/10.33549/physiolres.932383 |
| 23 | Wieckowski MR, Giorgi C, Lebiedzinska M, Duszynski J, Pinton P: Isolation of mitochondria-associated membranes and mitochondria from animal tissues and cells. Nature Protocols 2009;4(11):1582-90. DOI: 10.1038/nprot.2009.151.
https://doi.org/10.1038/nprot.2009.151 |
| 24 | Nemmar A, Al-Salam S, Beegam S, Zaaba NE, Elzaki O, Ali BH: Waterpipe smoke inhalation potentiates cardiac oxidative stress, inflammation, mitochondrial dysfunction, apoptosis and autophagy in experimental hypertension. Biomedicine & Pharmacotherapy 2023;158:114144. DOI: https://doi.org/10.1016/j.biopha.2022.114144.
https://doi.org/10.1016/j.biopha.2022.114144 |
| 25 | Spinazzi M, Casarin A, Pertegato V, Salviati L, Angelini C: Assessment of mitochondrial respiratory chain enzymatic activities on tissues and cultured cells. Nat Protoc 2012;7(6):1235-46. DOI: 10.1038/nprot.2012.058.
https://doi.org/10.1038/nprot.2012.058 |
| 26 | Olive PL, Banáth JP, Fjell CD: DNA strand breakage and DNA structure influence staining with propidium iodide using the alkaline comet assay. Cytometry 1994;16(4):305-12. DOI: https://doi.org/10.1002/cyto.990160404.
https://doi.org/10.1002/cyto.990160404 |
| 27 | Nemmar A, Al-Salam S, Beegam S, Yuvaraju P, Ali BH: Gum Arabic Ameliorates Impaired Coagulation and Cardiotoxicity Induced by Water-Pipe Smoke Exposure in Mice. Front Physiol 2019;10:53. DOI: 10.3389/fphys.2019.00053.
https://doi.org/10.3389/fphys.2019.00053 |
| 28 | Nemmar A, Al-Salam S, Yuvaraju P, Beegam S, Yasin J, Ali BH: Chronic exposure to water-pipe smoke induces cardiovascular dysfunction in mice. Am J Physiol Heart Circ Physiol 2017;312(2):H329-H39. DOI: 10.1152/ajpheart.00450.2016.
https://doi.org/10.1152/ajpheart.00450.2016 |
| 29 | Hu J, Wu Y, Wang Z, Li Z, Zhou Y, Wang H, Bao X, Hao J: Real-world fuel efficiency and exhaust emissions of light-duty diesel vehicles and their correlation with road conditions. J Environ Sci (China) 2012;24(5):865-74. DOI: 10.1016/s1001-0742(11)60878-4.
https://doi.org/10.1016/S1001-0742(11)60878-4 |
| 30 | Alexander M, Eissa M, McDermott-Levy R, Osborne R, Pleuss E, Prabhakaran P, Sorensen C: COP26: Looking forward from Glasgow by placing health at the center of climate action. The Journal of Climate Change and Health 2022;5:100117. DOI: https://doi.org/10.1016/j.joclim.2022.100117.
https://doi.org/10.1016/j.joclim.2022.100117 |
| 31 | Loussouarn M, Krieger-Liszkay A, Svilar L, Bily A, Birtić S, Havaux M: Carnosic Acid and Carnosol, Two Major Antioxidants of Rosemary, Act through Different Mechanisms. Plant Physiol 2017;175(3):1381-94. DOI: 10.1104/pp.17.01183.
https://doi.org/10.1104/pp.17.01183 |
| 32 | Cai P, Yan S, Lu Y, Zhou X, Wang X, Wang M, Yin Z: Carnosol inhibits osteoclastogenesis in vivo and in vitro by blocking the RANKL‑induced NF‑κB signaling pathway. Mol Med Rep 2022;26(1). DOI: 10.3892/mmr.2022.12741.
https://doi.org/10.3892/mmr.2022.12741 |
| 33 | Gonçalves C, Fernandes D, Silva I, Mateus V: Potential Anti-Inflammatory Effect of Rosmarinus officinalis in Preclinical In vivo Models of Inflammation. Molecules 2022;27(3). DOI: 10.3390/molecules27030609.
https://doi.org/10.3390/molecules27030609 |
| 34 | Lin C, Zhang X, Su Z, Xiao J, Lv M, Cao Y, Chen Y: Carnosol Improved Lifespan and Healthspan by Promoting Antioxidant Capacity in Caenorhabditis elegans. Oxid Med Cell Longev 2019;2019:5958043. DOI: 10.1155/2019/5958043.
https://doi.org/10.1155/2019/5958043 |
| 35 | Loo WY, Yue Y, Fan CB, Bai LJ, Dou YD, Wang M, Liang H, Cheung MN, Chow LW, Li JL, Tian Y, Qing L: Comparing serum levels of cardiac biomarkers in cancer patients receiving chemotherapy and subjects with chronic periodontitis. J Transl Med 2012;10 Suppl 1(Suppl 1):S5. DOI: 10.1186/1479-5876-10-s1-s5.
https://doi.org/10.1186/1479-5876-10-S1-S5 |
| 36 | Vuolteenaho O, Ala-Kopsala M, Ruskoaho H: BNP as a biomarker in heart disease. Adv Clin Chem 2005;40:1-36.
https://doi.org/10.1016/S0065-2423(05)40001-3 |
| 37 | Ramadi KB, Mohamed YA, Al-Sbiei A, Almarzooqi S, Bashir G, Al DA, Sarawathiamma D, Qadri S, Yasin J, Nemmar A, Fernandez-Cabezudo MJ, Haik Y, Al-Ramadi BK: Acute systemic exposure to silver-based nanoparticles induces hepatotoxicity and NLRP3-dependent inflammation. Nanotoxicology 2016:1-14.
https://doi.org/10.3109/17435390.2016.1163743 |
| 38 | Zaaba NE, Al-Salam S, Beegam S, Elzaki O, Yasin J, Nemmar A: Catalpol Attenuates Oxidative Stress and Inflammation via Mechanisms Involving Sirtuin-1 Activation and NF-κB Inhibition in Experimentally-Induced Chronic Kidney Disease. Nutrients 2023;15(1). DOI: 10.3390/nu15010237.
https://doi.org/10.3390/nu15010237 |
| 39 | Al Za'abi M, Al Busaidi M, Yasin J, Schupp N, Nemmar A, Ali BH: Development of a new model for the induction of chronic kidney disease via intraperitoneal adenine administration, and the effect of treatment with gum acacia thereon. Am J Transl Res 2015;7(1):28-38.
https://doi.org/10.1096/fasebj.29.1_supplement.938.3 |
| 40 | Mukhopadhyay P, Eid N, Abdelmegeed MA, Sen A: Interplay of Oxidative Stress, Inflammation, and Autophagy: Their Role in Tissue Injury of the Heart, Liver, and Kidney. Oxid Med Cell Longev 2018;2018:2090813. DOI: 10.1155/2018/2090813.
https://doi.org/10.1155/2018/2090813 |
| 41 | Savitskaya MA, Onishchenko GE: Mechanisms of apoptosis. Biochemistry (Moscow) 2015;80(11):1393-405. DOI: 10.1134/S0006297915110012.
https://doi.org/10.1134/S0006297915110012 |
| 42 | Li Z, Jo J, Jia J-M, Lo S-C, Whitcomb DJ, Jiao S, Cho K, Sheng M: Caspase-3 Activation via Mitochondria Is Required for Long-Term Depression and AMPA Receptor Internalization. Cell 2010;141(5):859-71. DOI: https://doi.org/10.1016/j.cell.2010.03.053.
https://doi.org/10.1016/j.cell.2010.03.053 |
| 43 | Boovarahan SR, Kurian GA: Mitochondrial dysfunction: a key player in the pathogenesis of cardiovascular diseases linked to air pollution. Rev Environ Health 2018;33(2):111-22. DOI: 10.1515/reveh-2017-0025.
https://doi.org/10.1515/reveh-2017-0025 |
| 44 | Geng Y, Wang Y, Sun R, Kang X, Zhao H, Zhu M, Sun Y, Hu Y, Wang Z, Tian X, Zhao Y, Yao J: Carnosol alleviates nonalcoholic fatty liver disease by inhibiting mitochondrial dysfunction and apoptosis through targeting of PRDX3. Toxicol Appl Pharmacol 2021;432:115758. DOI: 10.1016/j.taap.2021.115758.
https://doi.org/10.1016/j.taap.2021.115758 |
| 45 | Chen Y, Qin Q, Zhao W, Luo D, Huang Y, Liu G, Kuang Y, Cao Y, Chen Y: Carnosol Reduced Pathogenic Protein Aggregation and Cognitive Impairment in Neurodegenerative Diseases Models via Improving Proteostasis and Ameliorating Mitochondrial Disorders. Journal of Agricultural and Food Chemistry 2022;70(34):10490-505. DOI: 10.1021/acs.jafc.2c02665.
https://doi.org/10.1021/acs.jafc.2c02665 |
| 46 | Muslin AJ: MAPK signalling in cardiovascular health and disease: molecular mechanisms and therapeutic targets. Clin Sci (Lond) 2008;115(7):203-18. DOI: 10.1042/CS20070430.
https://doi.org/10.1042/CS20070430 |
| 47 | Pashkow FJ, Watumull DG, Campbell CL: Astaxanthin: A Novel Potential Treatment for Oxidative Stress and Inflammation in Cardiovascular Disease. The American Journal of Cardiology 2008;101(10, Supplement):S58-S68. DOI: https://doi.org/10.1016/j.amjcard.2008.02.010.
https://doi.org/10.1016/j.amjcard.2008.02.010 |
| 48 | Liu T, Zhang L, Joo D, Sun SC: NF-κB signaling in inflammation. Signal Transduct Target Ther 2017;2:17023-. DOI: 10.1038/sigtrans.2017.23.
https://doi.org/10.1038/sigtrans.2017.23 |
| 49 | Li L, Pan Z, Ning D, Fu Y: Rosmanol and Carnosol Synergistically Alleviate Rheumatoid Arthritis through Inhibiting TLR4/NF-κB/MAPK Pathway. Molecules 2021;27(1). DOI: 10.3390/molecules27010078.
https://doi.org/10.3390/molecules27010078 |
| 50 | Lo A-H, Liang Y-C, Lin-Shiau S-Y, Ho C-T, Lin J-K: Carnosol, an antioxidant in rosemary, suppresses inducible nitric oxide synthase through down-regulating nuclear factor-κB in mouse macrophages. Carcinogenesis 2002;23(6):983-91. DOI: 10.1093/carcin/23.6.983.
https://doi.org/10.1093/carcin/23.6.983 |
| 51 | Wang L-C, Wei W-H, Zhang X-W, Liu D, Zeng K-W, Tu P-F: An Integrated Proteomics and Bioinformatics Approach Reveals the Anti-inflammatory Mechanism of Carnosic Acid. Frontiers in Pharmacology 2018;9.
https://doi.org/10.3389/fphar.2018.00370 |
| 52 | Li G, Wu H, Sun L, Cheng K, Lv Z, Chen K, Qian F, Li Y: (-)-α-Bisabolol Alleviates Atopic Dermatitis by Inhibiting MAPK and NF-κB Signaling in Mast Cell. Molecules 2022;27(13). DOI: 10.3390/molecules27133985.
https://doi.org/10.3390/molecules27133985 |
| 53 | Ministrini S, Puspitasari YM, Beer G, Liberale L, Montecucco F, Camici GG: Sirtuin 1 in Endothelial Dysfunction and Cardiovascular Aging. Front Physiol 2021;12:733696. DOI: 10.3389/fphys.2021.733696.
https://doi.org/10.3389/fphys.2021.733696 |
| 54 | Pan W, Yu H, Huang S, Zhu P: Resveratrol Protects against TNF-α-Induced Injury in Human Umbilical Endothelial Cells through Promoting Sirtuin-1-Induced Repression of NF-KB and p38 MAPK. PLoS One 2016;11(1):e0147034. DOI: 10.1371/journal.pone.0147034.
https://doi.org/10.1371/journal.pone.0147034 |
| 55 | Kauppinen A, Suuronen T, Ojala J, Kaarniranta K, Salminen A: Antagonistic crosstalk between NF-κB and SIRT1 in the regulation of inflammation and metabolic disorders. Cellular Signalling 2013;25(10):1939-48. DOI: https://doi.org/10.1016/j.cellsig.2013.06.007.
https://doi.org/10.1016/j.cellsig.2013.06.007 |
| 56 | Teertam SK, Prakash Babu P: Differential role of SIRT1/MAPK pathway during cerebral ischemia in rats and humans. Scientific Reports 2021;11(1):6339. DOI: 10.1038/s41598-021-85577-9.
https://doi.org/10.1038/s41598-021-85577-9 |
| 57 | Zhao H, Wang Z, Tang F, Zhao Y, Feng D, Li Y, Hu Y, Wang C, Zhou J, Tian X, Yao J: Carnosol-mediated Sirtuin 1 activation inhibits Enhancer of Zeste Homolog 2 to attenuate liver fibrosis. Pharmacol Res 2018;128:327-37. DOI: 10.1016/j.phrs.2017.10.013.
https://doi.org/10.1016/j.phrs.2017.10.013 |
| 58 | Teng L, Fan L, Peng Y, He X, Chen H, Duan H, Yang F, Lin D, Lin Z, Li H, Shao B: Carnosic Acid Mitigates Early Brain Injury After Subarachnoid Hemorrhage: Possible Involvement of the SIRT1/p66shc Signaling Pathway. Front Neurosci 2019;13:26. DOI: 10.3389/fnins.2019.00026.
https://doi.org/10.3389/fnins.2019.00026 |