Prazosin Potentiates Mast Cell-Stabilizing Property of Adrenaline
Keywords
Abstract
Background/Aims:
Adrenaline quickly inhibits the release of histamine from mast cells. Besides β2-adrenergic receptors, several in vitro studies also indicate the involvement of α-adrenergic receptors in the process of exocytosis. Since exocytosis in mast cells can be detected electrophysiologically by the changes in the membrane capacitance (Cm), its continuous monitoring in the presence of drugs would determine their mast cell-stabilizing properties.Methods:
Employing the whole-cell patch-clamp technique in rat peritoneal mast cells, we examined the effects of adrenaline on the degranulation of mast cells and the increase in the Cm during exocytosis. We also examined the degranulation of mast cells in the presence or absence of α-adrenergic receptor agonists or antagonists.Results:
Adrenaline dose-dependently suppressed the GTP-γ-S-induced increase in the Cm and inhibited the degranulation from mast cells, which was almost completely erased in the presence of butoxamine, a β2-adrenergic receptor antagonist. Among α-adrenergic receptor agonists or antagonists, high dose prazosin, a selective α1-adrenergic receptor antagonist, significantly reduced the ratio of degranulating mast cells and suppressed the increase in the Cm.Conclusion:
This study provided electrophysiological evidence for the first time that adrenaline dose-dependently inhibited the process of exocytosis, confirming its usefulness as a potent mast cell-stabilizer. The pharmacological blockade of α1-adrenergic receptor by prazosin synergistically potentiated such mast cell-stabilizing property of adrenaline, which is primarily mediated by β2-adrenergic receptors.Introduction
Anaphylaxis is a severe allergic reaction and a potentially life-threatening acute multisystem syndrome caused by the sudden release of mast cell-derived mediators [1]. In the treatment, adrenaline, a non-specific adrenergic receptor agonist, is the first-choice drug, since it immediately suppresses further release of chemical mediators from mast cells [2]. Concerning the mechanisms, β2-adrenergic receptors are considered to be primarily responsible, because the stimulation of these receptors strongly inhibits FcεRI (high affinity receptors for IgE)-dependent calcium mobilization in the cells [3]. Previously, several in vitro studies also demonstrated the presence of α-adrenergic receptors in mast cells [4] and indicated their involvement in the activation of the cells [5, 6]. Based on these findings, later in vivo studies actually showed the therapeutic efficacy of prazosin, a specific α1-adrenergic receptor antagonist, for the histamine-induced bronchoconstriction in patients with asthma [7, 8]. To determine the effects of adrenaline or α-adrenergic receptor agonists/antagonists on the stabilization of mast cells, previous in vitro studies measured the drug-induced changes in histamine release from mast cells [6, 9-11]. However, they were not enough to monitor the whole process of exocytosis, since mast cells also release fibrogenic factors, growth factors and inflammatory cytokines in addition to chemical mediators [12]. In our series of patch-clamp studies, by detecting the changes in whole-cell membrane capacitance (Cm) in mast cells, we provided electrophysiological evidence that anti-allergic drugs, anti-microbial drugs, corticosteroids and food constituents inhibit the process of exocytosis, and thus exert mast cell-stabilizing properties [13-17]. In the present study, employing the same standard patch-clamp whole-cell recording technique in rat peritoneal mast cells, we examined the effects of adrenaline on the changes in the Cm to quantitatively determine its ability to stabilize mast cells. Additionally, we examined the effects of α-adrenergic receptor agonists or antagonists on the degranulation of mast cells to determine their involvement in the stabilization of mast cells. Here, this study provides electrophysiological evidence for the first time that adrenaline dose-dependently inhibits the process of exocytosis, confirming its usefulness as a potent mast cell-stabilizer. This study also shows that the pharmacological blockade of α1-adrenergic receptor by prazosin synergistically potentiates such mast cell-stabilizing property of adrenaline, which is primarily mediated by β2-adrenergic receptors.
Materials and Methods
Cell Sources and Preparation
Male Wistar rats no less than 25 weeks old were purchased from Japan SLC Inc. (Shizuoka, Japan). We profoundly anaesthetized the rats with isoflurane and sacrificed them by cervical dislocation. The protocols for the use of animals were approved by the Animal Care and Use Committee of Tohoku University Graduate School of Medicine and Miyagi University. As previously described [13-18], we washed rat peritoneum using standard external (bathing) solution which consists of (in mM): NaCl, 145; KCl, 4.0; CaCl2, 1.0; MgCl2, 2.0; HEPES, 5.0; bovine serum albumin, 0.01 % (pH 7.2 adjusted with NaOH) and isolated mast cells from the peritoneal cavity. We maintained the isolated mast cells at room temperature (22-24°C) to use within 8 hours. The suspension of mast cells was spread on a chamber placed on the headstage of an inverted microscope (Nikon, Tokyo, Japan). Mast cells were easy to distinguish from other cell types since they included characteristic secretory granules within the cells [13-16, 18].
Quantification of Mast Cell Degranulation
Adrenaline, purchased from Daiichi Sankyo, Inc. (Tokyo, Japan), dopamine, from Kyowa Hakko Kirin Co., Ltd. (Tokyo, Japan), phenylephrine hydrochloride, from Wako Pure Chem Ind. (Osaka, Japan), clonidine and yohimbine, from Tokyo Chemical Industry Co., Ltd. (Tokyo, Japan), were separately dissolved in the external solution at final concentrations of 1, 10, 100 μM and 1 mM. Prazosin hydrochloride, purchased from Tokyo Chemical Industry Co., Ltd. was dissolved at final concentrations of 0.01, 0.1, 1 μM. Butoxamine hydrochloride, purchased from Sigma Aldrich Co. (St. Louis, MO, USA), or prazosin was also dissolved in the external solution containing 1 mM adrenaline at the final concentrations of 1 mM or 1 μM, respectively. After we incubated mast cells in these solutions or a solution without the reagents, exocytosis was externally induced by compound 48/80 (Sigma-Aldrich; final concentration 10 μg/ml) [13-18]. We obtained bright-field images from randomly chosen 0.1-mm2 fields of view (10 views from each condition), as we described previously [13-18]. We counted degranulated mast cells (definition; cells surrounded by more than 8 granules outside the cell membrane) and calculated their ratio to all mast cells.
Electrical Setup and Membrane Capacitance Measurements
As we described in our previous studies [13-18], we employed an EPC-9 patch-clamp amplifier system (HEKA Electronics, Lambrecht, Germany) and conducted standard whole-cell patch-clamp recordings. Briefly, we maintained the patch pipette resistance between 4-6 MΩ when plugged with internal (patch pipette) solution which consists of (in mM): K-glutamate, 145; MgCl2, 2.0; Hepes, 5.0 (pH 7.2 adjusted with KOH). We added 100 μM guanosine 5’-o-(3-thiotriphosphate) (GTP-γ-S) (EMD Bioscience Inc., La Jolla, CA, USA) into the internal solution to endogenously induce exocytosis in mast cells [13-18]. We induced a giga-seal formation on a single mast cell spread in the external solutions containing no drug, different concentrations of adrenaline or dopamine (1, 10, 100 μM and 1 mM). Then we briefly sucked the pipette to rupture the patch membrane and perfused GTP-γ-S into the cells. We maintained the series resistance below 10 MΩ during the whole-cell recordings. To monitor the membrane capacitance of mast cells, we conducted a sine plus DC protocol employing the Lock-in amplifier of an EPC-9 Pulse program. We superimposed an 800-Hz sinusoidal command voltage on the holding potential of -80 mV. We continuously monitored the membrane capacitance (Cm), membrane conductance (Gm) and series conductance (Gs) during the whole-cell recording configuration. We performed all experiments at room temperature.
Statistical Analyses
Data were analyzed using PulseFit software (HEKA Electronics, Lambrecht, Germany) and Microsoft Excel (Microsoft Corporation, Redmond, Wash., USA) and reported as means ± SEM. Statistical significance was assessed by two-way ANOVA. A value of p < 0.05 was considered significant.
Results
Effects of adrenaline and dopamine on degranulation of rat peritoneal mast cells
Mast cells incubated in the external solution with compound 48/80 (10 μg/ml) showed more wrinkles on their cell surface than those incubated without the compound (Fig. 1Ab vs. a). They released more secretory granules due to exocytosis (Fig. 1Ab). Mast cells that were pre-incubated with relatively lower doses of adrenaline, a non-selective agonist of adrenergic receptors (1, 10 μM; Fig. 1Ac, d), showed similar findings to those that were incubated in the external solution alone (Fig. 1Ab). However, mast cells pre-incubated with relatively higher doses of adrenaline (100 μM, 1 mM; Fig. 1Ae, f) did not show such findings characteristic to exocytosis. On the other hand, almost all mast cells that were pre-incubated with dopamine, a non-selective agonist of dopamine receptors (1, 10, 100 μM and 1 mM), showed typical findings of exocytosis regardless of their concentrations (Fig. 1Ag to j).
To quantitatively determine such effects of adrenaline and dopamine on exocytosis, we then counted the numbers of degranulating mast cells and calculated their ratio to all mast cells (Fig. 1B and C). In the absence of adrenaline, compound 48/80 caused degranulation in 80.0 ± 1.4 % of the entire mast cells (n=10; Fig. 1B). Relatively lower concentrations of adrenaline (1, 10 μM) significantly decreased the number of degranulating mast cells dose-dependently (1 μM, 63.9 ± 2.3 %, n=15, p < 0.05; 10 μM, 56.7 ± 5.4 %, n=14, p < 0.05; Fig. 1B). Additionally, with higher concentrations (100 μM, 1 mM), adrenaline markedly reduced the numbers of degranulating mast cells (100 μM, 32.9 ± 2.1 %, n=14, p < 0.05; 1 mM, 24.1 ± 2.3 %, n=13, p < 0.05; Fig. 1B). Differing from adrenaline, dopamine did not significantly affect the numbers of degranulating mast cells regardless of their concentrations (Fig. 1C). From these results, consistent with the previous findings [9, 10], adrenaline, which suppresses the release of histamine, actually inhibited the degranulation of rat peritoneal mast cells dose-dependently.
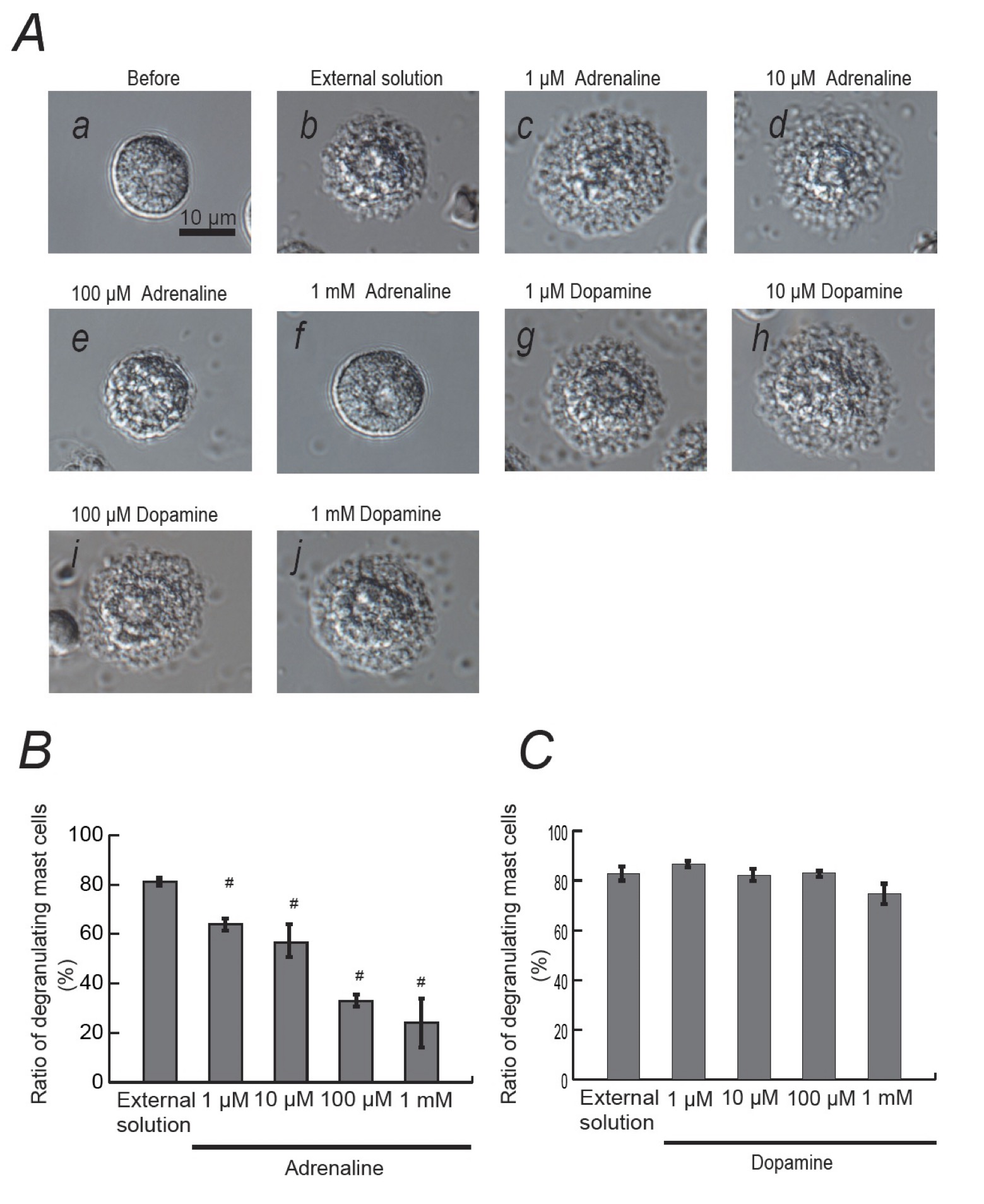
Fig. 1: Effects of adrenaline and dopamine on mast cell degranulation. A: Differential-interference contrast (DIC) microscopic images were taken before (a) and after exocytosis was externally induced by compound 48/80 in mast cells incubated in the external solutions containing no drug (b), 1 μM adrenaline (c), 10 μM adrenaline (d), 100 μM adrenaline (e), 1 mM adrenaline (f), 1 μM dopamine (g), 10 μM dopamine (h), 100 μM dopamine (i) and 1 mM dopamine (j). Effects of different concentrations (1, 10, 100 μM and 1 mM) of adrenaline (B) and dopamine (C). After the mast cells were incubated in the external solutions containing no drug or either drug, exocytosis was induced by compound 48/80. The numbers of degranulating mast cells were expressed as percentages of the total mast cell numbers in selected bright fields. # p < 0.05 vs. incubation in the external solution alone. Values are means ± SEM.
Effects of adrenaline and dopamine on whole-cell membrane capacitance in rat peritoneal mast cells
In our previous studies, microscopic changes in megakaryocyte or lymphocyte membranes were accurately monitored by measuring the whole-cell membrane capacitance (Cm) [19-26]. Of note, in mast cells, the process of degranulation during exocytosis was successively monitored by the increase in the Cm [13-18, 27, 28]. Hence, in our study, to quantitatively examine the effects of adrenaline or dopamine on the process of exocytosis, we pre-incubated mast cells in adrenaline- or dopamine- containing external solutions and measured the changes in Cm (Fig. 2 and 3). In these figures, we showed the effects of 1, 10, 100 μM and 1 mM adrenaline (Fig. 2) and dopamine (Fig. 3) on the Cm, Gs and Gm. Table 1 summarizes the changes in the Cm. Representing the endogenous induction of exocytosis [13-16, 18, 29, 30], the internal addition of GTP-γ-S into mast cells markedly increased the value of Cm (from 9.29 ± 0.37 to 34.0 ± 2.79 pF, n=9, p < 0.05; Table 1).
When mast cells were pre-incubated with lower doses of adrenaline (1, 10 μM), the addition of GTP-γ-S tended to increase the Cm similarly to that of mast cells pre-incubated with the external solution alone (Fig. 2A and B). However, compared to the external solution alone, the increase in the Cm (⊿Cm) was significantly suppressed (1 μM, 17.5 ± 6.83 pF, n=6, p < 0.05; 10 μM, 19.0 ± 2.03 pF, n=7, p < 0.05; Table 1). With higher doses (100 μM, 1 mM), adrenaline more markedly suppressed the GTP-γ-S-induced increase in the Cm (Fig. 2C and D; 100 μM, 7.61 ± 2.49 pF, n=8, p < 0.05; 1 mM, 5.41 ± 2.90 pF, n=6, p < 0.05; Table 1). In contrast, pre-incubation with dopamine did not significantly affect the the GTP-γ-S-induced increase in the Cm regardless of its concentrations (Fig. 3, Table 1). These results provided electrophysiological evidence for the first time that adrenaline inhibits the exocytotic process of mast cells dose-dependently. This strongly supported our findings that were obtained from Fig. 1.
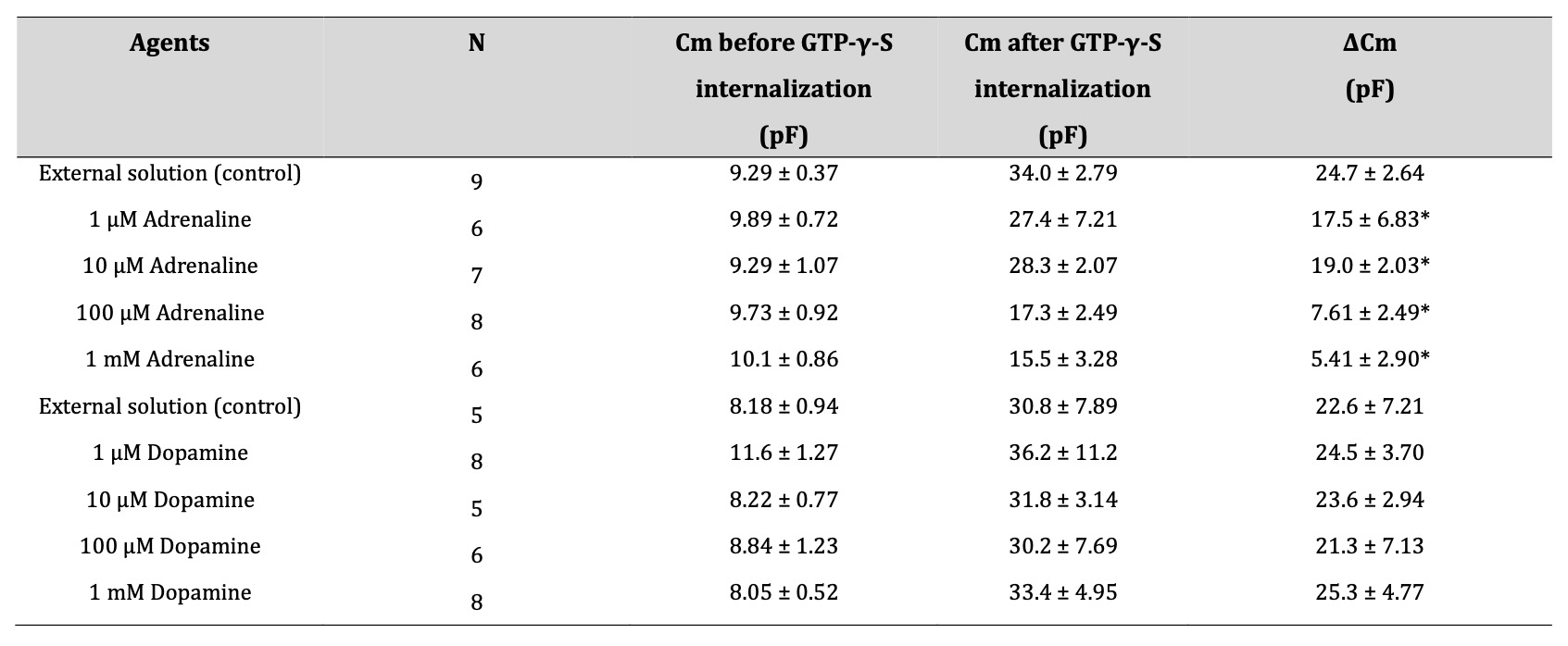
Table 1: Summary of changes in membrane capacitance. in external solutions containing adrenaline or dopamine. Values are means ± SEM. Cm = membrane capacitance. *p < 0.05 vs. ΔCm in external solution.
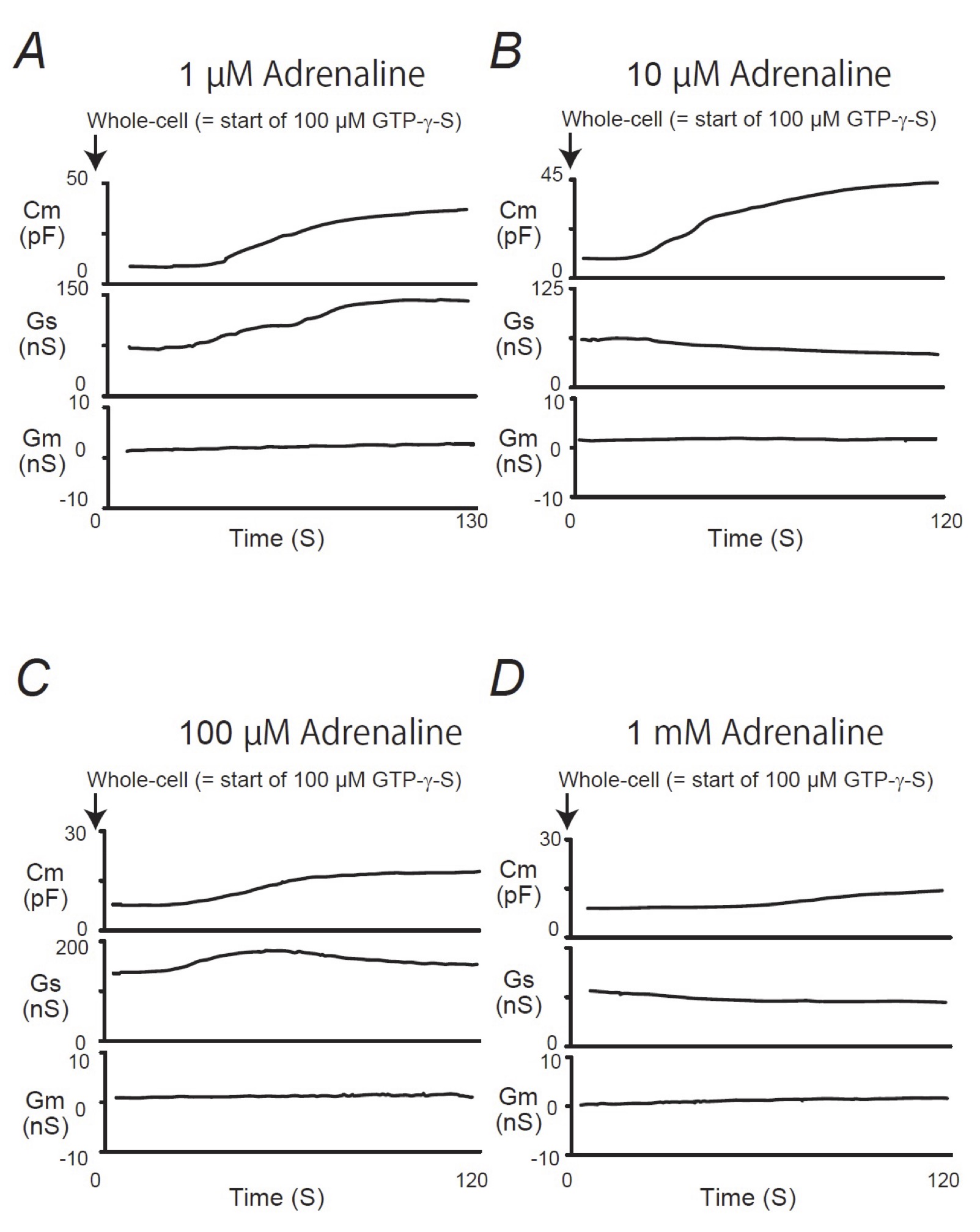
Fig. 2: Adrenaline-induced changes in mast cell membrane capacitance, series and membrane conductance during exocytosis. After the mast cells were incubated in the external solutions containing 1 μM (A), 10 μM (B), 100 μM (C) or 1mM adrenaline (D), the whole-cell recording configuration was established in single mast cells and dialysis with 100 μM GTP-γ-S was started. Membrane capacitance, series and membrane conductance were monitored for at least 90 sec. Cm, membrane capacitance; Gs, series conductance; Gm, membrane conductance.
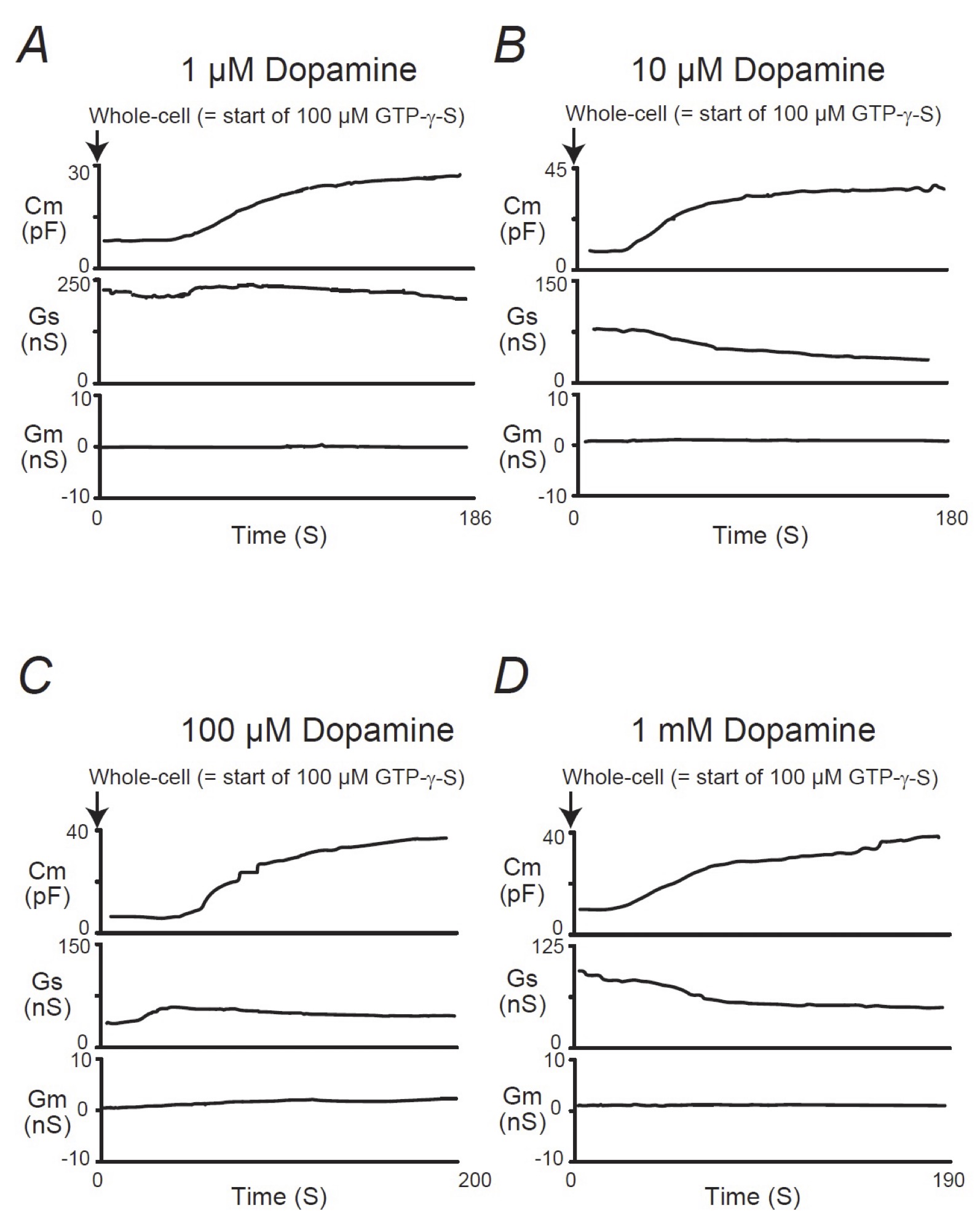
Fig. 3: Dopamine-induced changes in mast cell membrane capacitance, series and membrane conductance during exocytosis. After the mast cells were incubated in the external solutions containing 1 μM (A), 10 μM (B), 100 μM (C) or 1mM dopamine (D), the whole-cell recording configuration was established in single mast cells and dialysis with 100 μM GTP-γ-S was started. Membrane capacitance, series and membrane conductance were monitored for at least 90 sec. Cm, membrane capacitance; Gs, series conductance; Gm, membrane conductance.
Effects of β2-adrenergic receptor antagonist on adrenaline-induced inhibition of mast cell degranulation
Mast cells express numerous receptors on their cell surface that transduce stimulatory or inhibitory signals for degranulation [31, 32]. Among them, β2-adrenergic receptor is the major one that transduces inhibitory signals for exocytosis [3]. Since adrenaline is one of the most potent non-specific stimulators of adrenergic receptors, we examined the involvement of this receptor-mediated pathway in the adrenaline-induced inhibition of exocytosis. Consistent with our findings obtained from Fig. 1A and B, pre-incubation with 1mM adrenaline halted the induction of exocytosis in mast cells (Fig. 4Ab vs. a) by markedly suppressing the numbers of degranulating cells (Fig. 4B). However, in the presence of 1 mM butoxamine, a specific β2-adrenergic receptor antagonist, such inhibitory effect of adrenaline on exocytosis was almost completely erased (Fig. 4Ac and 4B). These results confirmed the previous findings in rat peritoneal mast cells that the stimulation of β2-adrenergic receptors, which is linked to a cyclic AMP-dependent calcium mobilization via the coupling of G-proteins [33], is the major pathway for the adrenaline-induced inhibition of exocytosis [3].
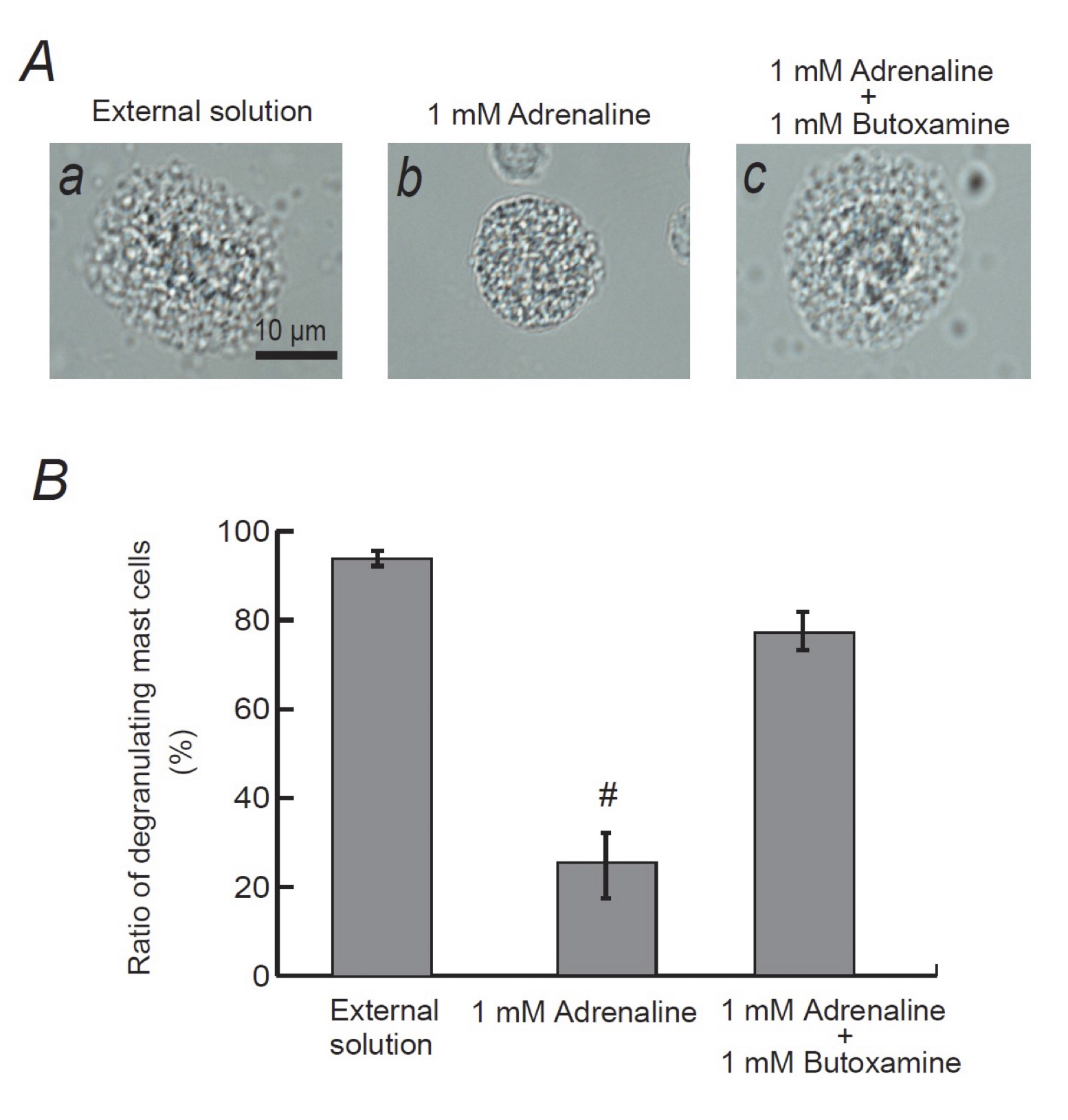
Fig. 4: Effects of β2-adrenergic receptor antagonist on adrenaline-induced inhibition of mast cell degranulation. A: Differential-interference contrast (DIC) microscopic images were taken after exocytosis was externally induced by compound 48/80 in mast cells incubated in the external solutions containing no drug (a), 1 mM adrenaline (b) or 1 mM adrenaline in the presence of 1 mM butoxamine (c). (B) After exocytosis was induced in mast cells incubated in the external solutions containing no drug, 1 mM adrenaline with or without the presence of 1 mM butoxamine, the numbers of degranulating mast cells were expressed as percentages of the total mast cell numbers in selected bright fields. # p<0.05 vs. incubation in the external solution alone. Values are means ± SEM.
Involvement of α-adrenergic receptors in degranulation of rat peritoneal mast cells
In addition to β2-adrenergic receptors that transduce inhibitory signals for the degranulation of mast cells (Fig. 4), studies revealed the localization of α1-adrenergic receptors on mast cell membranes [4] and also provided in vivo evidence for the presence of α2-adrenergic receptors [11, 34]. To reveal the involvement of these adrenergic receptors in the degranulation of mast cells, we examined the effects of the receptor agonists or antagonists.
(I) Effects of α1- or α2-adrenergic receptor “agonists” on degranulation of rat peritoneal mast cells
Consistent with our results shown in Fig. 1B and C, compound 48/80 caused degranulation in 80.0 ± 1.4 % of the entire mast cells in the external solution alone (n=10; Fig. 5A). However, pre-incubation with 1, 10, 100 μM and 1 mM phenylephrine, a selective α1-adrenergic receptor agonist, did not significantly affect the numbers of degranulating mast cells regardless of their concentrations (Fig. 5A). Similarly, pre-incubation with different concentrations of clonidine, a selective α2-adrenergic receptor agonist, did not alter the ratio of degranulating mast cells, either (Fig. 5B).
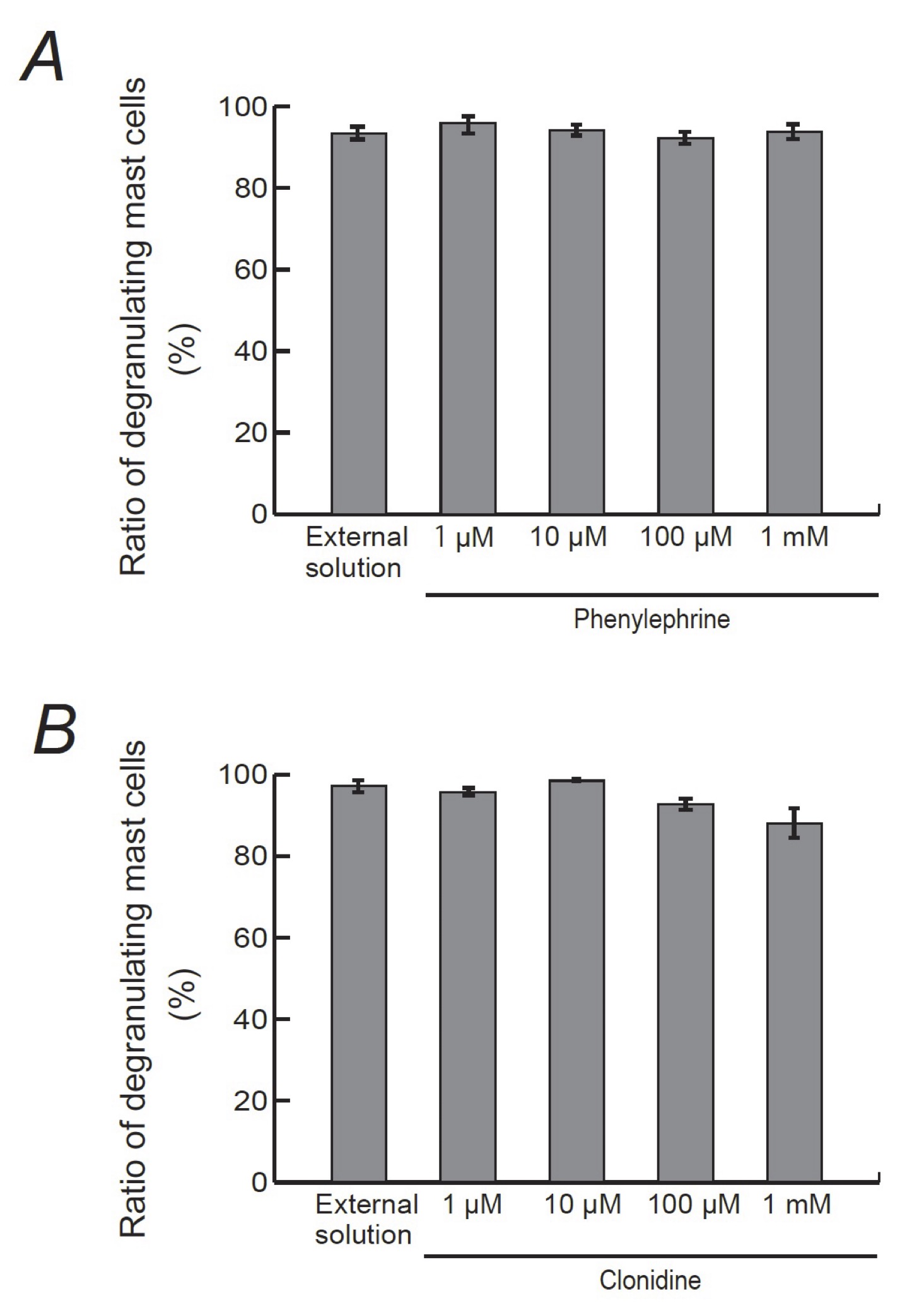
Fig. 5: Effects of α1- or α2-adrenergic receptor agonists on mast cell degranulation. Effects of different concentrations (1, 10, 100 μM and 1 mM) of phenylephrine (A) and clonidine (B). After the mast cells were incubated in the external solutions containing no drug or either drug, exocytosis was induced by compound 48/80. The numbers of degranulating mast cells were expressed as percentages of the total mast cell numbers in selected bright fields. Values are means ± SEM.
(II) Effects of α1- or α2-adrenergic receptor “antagonists” on exocytosis of rat peritoneal mast cells
Since α1- and α2-adrenergic receptor agonists did not affect the process of exocytosis in mast cells (Fig. 5), we then examined the effects of α1- and α2-adrenergic receptor antagonists (Fig. 6). The physiological concentration of prazosin, a selective α1-adrenergic receptor antagonist, is as low as between 2.60 and 26.0 nM in humans [35], which is by far lower than that of adrenaline, dopamine, phenylephrine and clonidine [36]. Additionally, in some in vitro studies, prazosin with concentrations as low as 0.1 μM was enough to exert inhibitory effects on the α1-adrenergic receptor-mediated proliferation in cultured vascular smooth muscle cells [37]. Therefore, in the present study, we tried doses from as low as 0.01 up to 1 μM (Fig. 6A). Relatively lower doses, such as 0.01 and 0.1 μM, did not significantly affect the numbers of degranulating mast cells (Fig. 6Aa). However, 1 μM prazosin alone significantly reduced the ratio of degranulating mast cells compared to that of the external solution (from 84.5 ± 2.1 % to 64.7 ± 3.8 %, n=10, p < 0.05). In mast cells, the process of degranulation during exocytosis was monitored by the increase in the Cm [13-18, 27, 28]. Actually, in the present study, the ratio of degraunalting mast cells was well correlated with the GTP-γ-S-induced increase in the Cm (⊿Cm) (Fig. 1 to 3, Table 1). Therefore, we additionally examined the effects of prazosin on the ⊿Cm (Fig. 6Ab). Similarly to the ratio of degranulating mast cells (Fig. 6Aa), low dose prazosin did not significantly affect the ⊿Cm (Fig. 6Ab). However, 1 μM prazosin alone significantly decreased the ⊿Cm compared to that of the external solution (from 19.6 ± 2.38 pF to 10.4 ± 1.68 pF, n=6, p < 0.05; Fig. 6Ab). These results provided electrophysiological evidence that high dose prazosin can inhibit the process of exocytosis in mast cells. In contrast, however, yohimbine, a selective α2-adrenergic receptor antagonist, did not affect the ratio of degranulating mast cells (Fig. 6B). These results suggested that the process of exocytosis in mast cells may be partially mediated by α1-adrenergic receptors, but not by α2-adrenergic receptors.
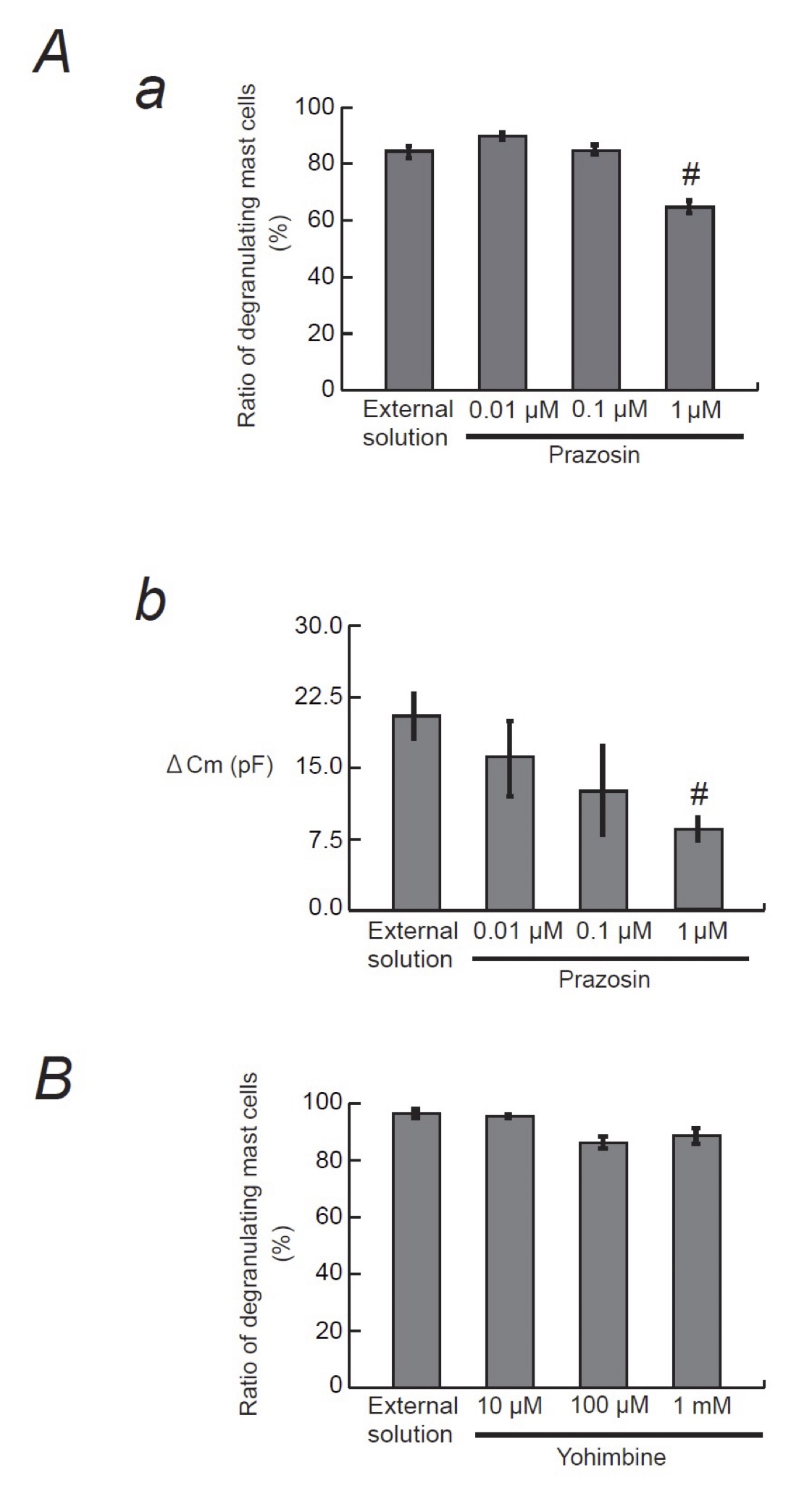
Fig. 6: Effects of α1- or α2-adrenergic receptor antagonists on mast cell degranulation. A: Effects of prazosin on mast cell degranulation and membrane capacitance. a: After the mast cells were incubated in the external solutions containing no drug or different concentrations (0.01, 0.1 and 1 μM) of prazosin, exocytosis was induced by compound 48/80. The numbers of degranulating mast cells were expressed as percentages of the total mast cell numbers in selected bright fields. b: After the mast cells were incubated in the external solutions containing no drug or different concentrations (0.01, 0.1 and 1 μM) of prazosin, the whole-cell recording configuration was established in single mast cells and dialysis with 100 μM GTP-γ-S was started. The GTP-γ-S-induced increase in the Cm (⊿Cm) was calculated. B: Effects of yohimbine on mast cell degranulation. After the mast cells were incubated in the external solutions containing no drug or different concentrations (10, 100 μM and 1 mM) of yohimbine, exocytosis was induced by compound 48/80. The numbers of degranulating mast cells were expressed as percentages of the total mast cell numbers in selected bright fields.# p<0.05 vs. incubation in the external solution alone. Values are means ± SEM.
Effects of prazosin on adrenaline-induced inhibition of mast cell degranulation
From our results, since 1 μM prazosin inhibited the process of exocytosis in mast cells (Fig. 6A), we finally examined its effect on the adrenaline-induced inhibition of exocytosis (Fig. 7). Consistent with our results shown Fig. 1A and B, pre-incubation with 1mM adrenaline halted the induction of exocytosis (Fig. 7Ab vs. a) and markedly reduced the numbers of degranulating mast cells (Fig. 7B). In the presence of 1 μM prazosin, such inhibitory effect of adrenaline on exocytosis was augmented (Fig. 7B) and the induction of exocytosis was almost totally suppressed (Fig. 7Ac). These results suggested that the blockade of α1-adrenergic receptors by prazosin can synergistically potentiate the β2-adrenergic receptor-mediated inhibition of exocytosis in mast cells.
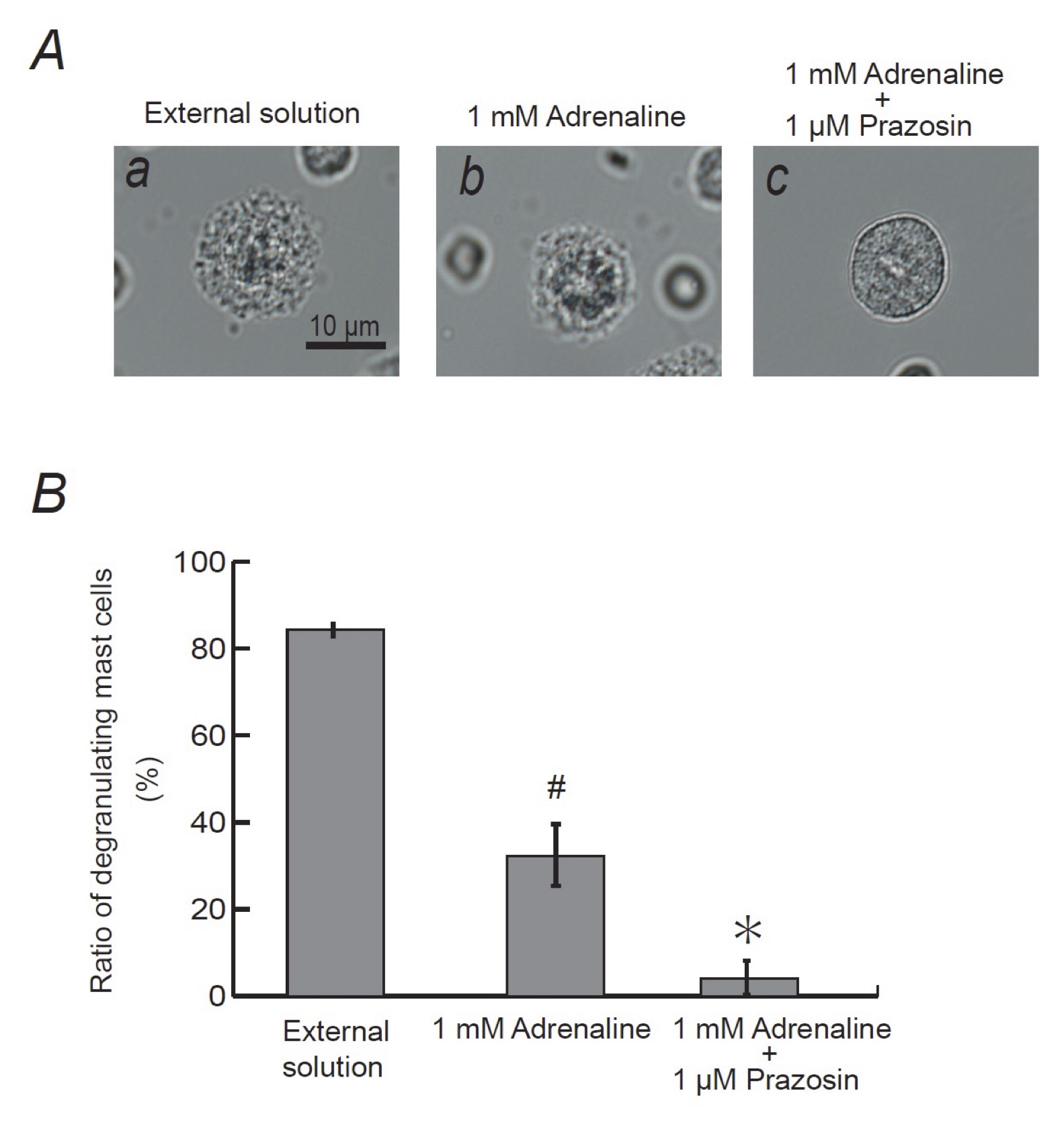
Fig. 7: Effects of α1-adrenergic receptor antagonist on adrenaline-induced inhibition of mast cell degranulation. A: Differential-interference contrast (DIC) microscopic images were taken after exocytosis was externally induced by compound 48/80 in mast cells incubated in the external solutions containing no drug (a), 1 mM adrenaline (b) or 1 mM adrenaline in the presence of 1 μM prazosin (c). (B) After exocytosis was induced in mast cells incubated in the external solutions containing no drug, 1 mM adrenaline with or without the presence of 1 μM prazosin, the numbers of degranulating mast cells were expressed as percentages of the total mast cell numbers in selected bright fields. # p<0.05 vs. incubation in the external solution alone. *p<0.05 vs. incubation in the external solution containing 1 mM adrenaline. Values are means ± SEM.
Discussion
For people experiencing anaphylaxis or those at risks of anaphylactic reaction, intramuscular injection of adrenaline, a non-selective agonist of β-adrenergic receptors, has been the first-choice of the treatment [2]. In previous studies, by measuring the amount of histamine released from mast cells, suppressive effects of adrenaline on the activation of mast cells were indirectly monitored [9, 10]. However, to precisely determine the ability of adrenaline on the stabilization of mast cells, the exocytotic process itself needs to be monitored, otherwise the release of all the chemical mediators or the inflammatory substances have to be evaluated. In our previous patch-clamp studies using rat peritoneal mast cells, the degranulating process during exocytosis was successively monitored by the gradual increase in the whole-cell Cm [15-18, 29, 38]. Employing this electrophysiological approach, our recent studies revealed the inhibitory effects of anti-allergic drugs, antibiotics, corticosteroids and food constituents, such as vitamins, on the exocytotic process of mast cells [13-17]. In these studies, the mast cell-stabilizing properties of the drugs were quantitatively determined by the suppressed value of Cm which is to be increased by the GTP-γ-S internalization [13, 14]. In the present study, applying the same approach, we provided direct evidence for the first time that adrenaline actually inhibits the process of exocytosis dose-dependently and thus exerts mast cell-stabilizing property.
The physiological concentration of adrenaline in the plasma is usually below 0.1 μM at the basal level [39, 40]. However, it reaches more than 0.3 μM up to 1.5 μM after intramuscular injection in the treatment of anaphylaxis [41, 42]. In the present study, 1 μM adrenaline significantly deceased the number of degranulating mast cells by approximately 20 % (Fig. 1B), which was consistent with the findings obtained from previous studies [9]. Additionally, we further revealed for the first time that adrenaline with higher doses, such as 10, 100 μM and 1 mM, more markedly suppressed the degranulation of mast cells dose-dependently (Fig. 1B). These findings could be clinically applied to the topical use of adrenaline on nasal mucosa [40], where higher concentrations are locally required before the drug is absorbed into the venous circulation by the transcellular diffusion [43]. In the present study, exocytosis was externally induced by compound 48/80 after pretreating mast cells with adrenaline. Similar to the antigen binding to IgE on mast cells that causes quick anaphylactic reaction, compound 48/80 initiates the degranulation of mast cells as immediately as 10 seconds after its addition [44]. Therefore, it would be difficult to examine the “therapeutic” effects of adrenaline on reversing the ongoing degranulation of mast cells. However, our study clearly demonstrated the “prophylactic” effects of adrenaline on suppressing the further initiation of exocytosis in mast cells. In our whole experiments, we used mast cells isolated from the peritoneal cavity of rats less than 25 weeks old, since mast cells isolated form these rats were viable enough to be easily induced exocytosis by the exogenous or endogenous pharmacological stimuli [13-17, 45].
In the present study, butoxamine, a β2-adrenergic receptor antagonist, almost totally restored the adrenaline-induced inhibition of mast cell degranulation (Fig. 4). This confirmed the previous findings that β2-adrenergic pathway is the major pathway in which adrenaline transduces inhibitory signals for the degranulation of mast cells [3, 33]. In addition to β2-adrenergic receptors, previous in vitro studies demonstrated the expression of α1-adrenergic receptors on mast cell membranes [4] or provided in vivo evidence indicating the presence of α2-adrenergic receptors [11, 34]. There are two types of mast cells that exist throughout the body [46]. One is the connective tissue type, which primarily exists in loose connective tissues, such as peritoneal cavity or skin. The other is the mucosal type, which primarily exists airway or gastrointestinal mucosa. In contrast to β2-adrenergic receptors that are expressed in both types of mast cells [47], α1-adrenergic receptors were shown to be expressed in mast cells isolated from heart connective tissue [4]. However, several in vitro studies using α-adrenergic agonists functionally demonstrated the presence of α-adrenergic receptors in mucosal type mast cells, such as human lung mast cells [5]. From our results, α2-adrenergic receptors were not likely to be involved in the process of exocytosis in mast cells, since both agonist and antagonist of the receptors did not affect the degranulation of mast cells (Fig. 5B and 6B). On the other hand, we noted for the first time that high dose prazosin, an α1-adrenergic receptor antagonist, significantly suppressed the degranulation of mast cells (Fig. 6A) and synergistically potentiated the adrenaline-induced inhibition of exocytosis (Fig. 7). In previous in vitro studies using human lung mast cells, stimulation of α1-adrenergic receptors increased the release of chemical mediators [5]. Based on this, later studies further demonstrated in humans that the pharmacological blockade of α1-adrenergic receptors actually ameliorated the airway hyperresponsiveness in patients with asthma [7, 8]. In this context, our results strongly suggested that the blockade of α1-adrenergic receptor by prazosin may also be useful in the treatment of anaphylaxis by potentiating the therapeutic efficacy of adrenaline. However, to exert such effects, prazosin with doses much higher than those of the physiological concentration was required (Fig. 6A), which can deteriorate hypotension due to the blockade of vascular α1-adrenergic receptors [48]. In such cases, the use of omalizumab or talizumab that directly inhibits the binding of IgE to FcεRI may be considered [49], since these reagents are more selective to immune systems compared to prazosin.
As we have shown in our patch-clamp studies, the elevation of the intracellular Ca2+ concentration ([Ca2+]i) primarily triggers exocytosis in mast cells [14]. According to previous studies using human lung mast cells, the elevation of the [Ca2+]i was primarily ascribable to the activity of Ca2+ activated K+ channels (Kca 3.1), because these channels facilitate the Ca2+ influx through store-operated calcium channels (SOCs) [50]. Upon activation, α1-adrenergic receptors stimulate phospholipase C (PLC) via the coupling of G proteins [51]. This enzymatically cleaves phosphatidylinositol triphosphate (PIP2) into inositol triphosphate (IP3) and diacylglycerol (DAG), which leads to the activation of protein kinase C (PKC) [52]. Since PKC is known to stimulate the activity of Kca 3.1 [53] or SOC, such as transient receptor potential canonical (TRPC) 1 and 6 [54, 55], the upstream blockade of α1-adrenergic receptor by prazosin, may inhibit the activity of these channels. Such induced decrease in [Ca2+]i was thought to be the mechanism by which prazosin exerts mast cell-stabilizing property. Alternatively, as we previously demonstrated in anti-allergic drugs or macrolide antibiotics [13, 14, 16], highly lipophilic prazosin [56], which is prone to penetrate into the plasma membrane and accumulate there, may have induced membrane stretch in mast cells. Such mechanical stimuli to the membranes would re-arrange the cytoskeletal structures, influencing the activity of the K+- or Ca2+-channels expressed in mast cells. Consequently, such induced changes in the [Ca2+]i were thought to contribute to the prazosin-induced inhibition of exocytosis.
Conclusion
In summary, this study provided electrophysiological evidence for the first time that adrenaline dose-dependently inhibits the process of exocytosis, confirming its usefulness as a potent mast cell-stabilizer. The pharmacological blockade of α1-adrenergic receptor by prazosin synergistically potentiated such mast cell-stabilizing property of adrenaline, which is primarily mediated by β2-adrenergic receptors.
Acknowledgements
NA performed the experiments and analyzed the data. IK designed the experiments, interpreted the results and wrote the paper. HT, YE, KS, TT and MY provided logistical support and advised on the project. All authors read and approved the final manuscript. This work was supported by the Salt Science Research Foundation, No. 2028, the Tojuro Iijima Foundation for Food Science and Technology, No. 2019-12, and Miyagi Kidney Foundation Grant to IK.
Disclosure Statement
The authors declare no conflicts of interest.
References
| 1 | Sampson HA, Munoz-Furlong A, Campbell RL, Adkinson NF, Jr., Bock SA, Branum A et al.: Second symposium on the definition and management of anaphylaxis: summary report--Second National Institute of Allergy and Infectious Disease/Food Allergy and Anaphylaxis Network symposium. J Allergy Clin Immunol 2006;117:391-397.
https://doi.org/10.1016/j.jaci.2005.12.1303 |
| 2 | Kemp SF, Lockey RF, Simons FE, World Allergy Organization ad hoc Committee on Epinephrine in A: Epinephrine: the drug of choice for anaphylaxis-a statement of the world allergy organization. World Allergy Organ J 2008;1:S18-26.
https://doi.org/10.1186/1939-4551-1-S2-S18 |
| 3 | Kuehn HS, Gilfillan AM: G protein-coupled receptors and the modification of FcepsilonRI-mediated mast cell activation. Immunol Lett 2007;113:59-69.
https://doi.org/10.1016/j.imlet.2007.08.007 |
| 4 | Schulze W, Fu ML: Localization of alpha 1-adrenoceptors in rat and human hearts by immunocytochemistry. Mol Cell Biochem 1996;163-164:159-165.
https://doi.org/10.1007/BF00408653 |
| 5 | Kaliner M, Orange RP, Austen KF: Immunological release of histamine and slow reacting substance of anaphylaxis from human lung. J Exp Med 1972;136:556-567.
https://doi.org/10.1084/jem.136.3.556 |
| 6 | Moroni F, Fantozzi R, Masini E, Mannaioni PF: The modulation of histamine release by alpha-adrenoceptors: evidences in murine neoplastic mast cells. Agents Actions 1977;7:57-61.
https://doi.org/10.1007/BF01964881 |
| 7 | Barnes PJ, Wilson NM, Vickers H: Prazosin, an alpha 1-adrenoceptor antagonist, partially inhibits exercise-induced asthma. J Allergy Clin Immunol 1981;68:411-415.
https://doi.org/10.1016/0091-6749(81)90193-7 |
| 8 | Jenkins C, Breslin AB, Marlin GE: The role of alpha and beta adrenoceptors in airway hyperresponsiveness to histamine. J Allergy Clin Immunol 1985;75:364-372.
https://doi.org/10.1016/0091-6749(85)90073-9 |
| 9 | Ng WH, Polosa R, Church MK: Adenosine bronchoconstriction in asthma: investigations into its possible mechanism of action. Br J Clin Pharmacol 1990;30 Suppl 1:89S-98S.
https://doi.org/10.1111/j.1365-2125.1990.tb05474.x |
| 10 | Graevskaya EE, Akhalaya MY, Goncharenko EN: Effects of cold stress and epinephrine on degranulation of peritoneal mast cells in rats. Bull Exp Biol Med 2001;131:333-335.
https://doi.org/10.1023/A:1017991817000 |
| 11 | Lindgren BR, Grundstrom N, Andersson RG: Comparison of the effects of clonidine and guanfacine on the histamine liberation from human mast cells and basophils and on the human bronchial smooth muscle activity. Arzneimittelforschung 1987;37:551-553.
|
| 12 | Gruber BL: Mast cells in the pathogenesis of fibrosis. Curr Rheumatol Rep 2003;5:147-153.
https://doi.org/10.1007/s11926-003-0043-3 |
| 13 | Baba A, Tachi M, Maruyama Y, Kazama I: Olopatadine inhibits exocytosis in rat peritoneal mast cells by counteracting membrane surface deformation. Cell Physiol Biochem 2015;35:386-396.
https://doi.org/10.1159/000369704 |
| 14 | Baba A, Tachi M, Ejima Y, Endo Y, Toyama H, Matsubara M et al.: Anti-Allergic Drugs Tranilast and Ketotifen Dose-dependently Exert Mast Cell-Stabilizing Properties. Cell Physiol Biochem 2016;38:15-27.
https://doi.org/10.1159/000438605 |
| 15 | Mori T, Abe N, Saito K, Toyama H, Endo Y, Ejima Y et al.: Hydrocortisone and dexamethasone dose-dependently stabilize mast cells derived from rat peritoneum. Pharmacol Rep 2016;68:1358-1365.
https://doi.org/10.1016/j.pharep.2016.09.005 |
| 16 | Kazama I, Saito K, Baba A, Mori T, Abe N, Endo Y et al.: Clarithromycin Dose-Dependently Stabilizes Rat Peritoneal Mast Cells. Chemotherapy 2016;61:295-303.
https://doi.org/10.1159/000445023 |
| 17 | Kazama I, Sato Y, Tamada T: Pyridoxine Synergistically Potentiates Mast Cell-Stabilizing Property of Ascorbic Acid. Cell Physiol Biochem 2022;56:282-292.
https://doi.org/10.33594/000000534 |
| 18 | Kazama I, Maruyama Y, Takahashi S, Kokumai T: Amphipaths differentially modulate membrane surface deformation in rat peritoneal mast cells during exocytosis. Cell Physiol Biochem 2013;31:592-600.
https://doi.org/10.1159/000350079 |
| 19 | Kazama I, Maruyama Y, Nakamichi S: Aspirin-induced microscopic surface changes stimulate thrombopoiesis in rat megakaryocytes. Clin Appl Thromb Hemost 2014;20:318-325.
https://doi.org/10.1177/1076029612461845 |
| 20 | Kazama I, Maruyama Y, Murata Y: Suppressive effects of nonsteroidal anti-inflammatory drugs diclofenac sodium, salicylate and indomethacin on delayed rectifier K+-channel currents in murine thymocytes. Immunopharmacol Immunotoxicol 2012;34:874-878.
https://doi.org/10.3109/08923973.2012.666249 |
| 21 | Kazama I, Maruyama Y, Matsubara M: Benidipine persistently inhibits delayed rectifier K(+)-channel currents in murine thymocytes. Immunopharmacol Immunotoxicol 2013;35:28-33.
https://doi.org/10.3109/08923973.2012.723011 |
| 22 | Kazama I, Maruyama Y: Differential effects of clarithromycin and azithromycin on delayed rectifier K(+)-channel currents in murine thymocytes. Pharm Biol 2013;51:760-765.
https://doi.org/10.3109/13880209.2013.764539 |
| 23 | Kazama I, Baba A, Maruyama Y: HMG-CoA reductase inhibitors pravastatin, lovastatin and simvastatin suppress delayed rectifier K(+)-channel currents in murine thymocytes. Pharmacol Rep 2014;66:712-717.
https://doi.org/10.1016/j.pharep.2014.03.002 |
| 24 | Baba A, Tachi M, Maruyama Y, Kazama I: Suppressive Effects of Diltiazem and Verapamil on Delayed Rectifier K+-Channel Currents in Murine Thymocytes. Pharmacol Rep 2015 67:959-964.
https://doi.org/10.1016/j.pharep.2015.01.009 |
| 25 | Kazama I, Ejima Y, Endo Y, Toyama H, Matsubara M, Baba A et al.: Chlorpromazine-induced changes in membrane micro-architecture inhibit thrombopoiesis in rat megakaryocytes. Biochim Biophys Acta 2015;1848:2805-2812.
https://doi.org/10.1016/j.bbamem.2015.08.013 |
| 26 | Kazama I, Baba A, Endo Y, Toyama H, Ejima Y, Matsubara M et al.: Salicylate Inhibits Thrombopoiesis in Rat Megakaryocytes by Changing the Membrane Micro-Architecture. Cell Physiol Biochem 2015;35:2371-2382.
https://doi.org/10.1159/000374039 |
| 27 | Fernandez JM, Neher E, Gomperts BD: Capacitance measurements reveal stepwise fusion events in degranulating mast cells. Nature 1984;312:453-455.
https://doi.org/10.1038/312453a0 |
| 28 | Lorenz D, Wiesner B, Zipper J, Winkler A, Krause E, Beyermann M et al.: Mechanism of peptide-induced mast cell degranulation. Translocation and patch-clamp studies. J Gen Physiol 1998;112:577-591.
https://doi.org/10.1085/jgp.112.5.577 |
| 29 | Neher E: The influence of intracellular calcium concentration on degranulation of dialysed mast cells from rat peritoneum. J Physiol 1988;395:193-214.
https://doi.org/10.1113/jphysiol.1988.sp016914 |
| 30 | Penner R, Neher E: Secretory responses of rat peritoneal mast cells to high intracellular calcium. FEBS Lett 1988;226:307-313.
https://doi.org/10.1016/0014-5793(88)81445-5 |
| 31 | McNeil BD, Pundir P, Meeker S, Han L, Undem BJ, Kulka M et al.: Identification of a mast-cell-specific receptor crucial for pseudo-allergic drug reactions. Nature 2015;519:237-241.
https://doi.org/10.1038/nature14022 |
| 32 | Katz HR: Inhibitory receptors and allergy. Curr Opin Immunol 2002;14:698-704.
https://doi.org/10.1016/S0952-7915(02)00400-4 |
| 33 | Chong LK, Morice AH, Yeo WW, Schleimer RP, Peachell PT: Functional desensitization of beta agonist responses in human lung mast cells. Am J Respir Cell Mol Biol 1995;13:540-546.
https://doi.org/10.1165/ajrcmb.13.5.7576689 |
| 34 | Lindgren BR, Brundin A, Andersson RG: Inhibitory effects of clonidine on the allergen-induced wheal-and-flare reactions in patients with extrinsic asthma. J Allergy Clin Immunol 1987;79:941-946.
https://doi.org/10.1016/0091-6749(87)90244-2 |
| 35 | Rubin PC, Butters L, Low RA, Reid JL: Clinical pharmacological studies with prazosin during pregnancy complicated by hypertension. Br J Clin Pharmacol 1983;16:543-547.
https://doi.org/10.1111/j.1365-2125.1983.tb02213.x |
| 36 | Vincent J, Meredith PA, Reid JL, Elliott HL, Rubin PC: Clinical pharmacokinetics of prazosin--1985. Clin Pharmacokinet 1985;10:144-154.
https://doi.org/10.2165/00003088-198510020-00002 |
| 37 | Yu SM, Tsai SY, Guh JH, Ko FN, Teng CM, Ou JT: Mechanism of catecholamine-induced proliferation of vascular smooth muscle cells. Circulation 1996;94:547-554.
https://doi.org/10.1161/01.CIR.94.3.547 |
| 38 | Penner R: Multiple signaling pathways control stimulus-secretion coupling in rat peritoneal mast cells. Proc Natl Acad Sci U S A 1988;85:9856-9860.
https://doi.org/10.1073/pnas.85.24.9856 |
| 39 | Body JJ, Cryer PE, Offord KP, Heath H, 3rd: Epinephrine is a hypophosphatemic hormone in man. Physiological effects of circulating epinephrine on plasma calcium, magnesium, phosphorus, parathyroid hormone, and calcitonin. J Clin Invest 1983;71:572-578.
https://doi.org/10.1172/JCI110802 |
| 40 | Sarmento Junior KM, Tomita S, Kos AO: Topical use of adrenaline in different concentrations for endoscopic sinus surgery. Braz J Otorhinolaryngol 2009;75:280-289.
https://doi.org/10.1016/S1808-8694(15)30791-6 |
| 41 | Simons FE, Gu X, Simons KJ: Epinephrine absorption in adults: intramuscular versus subcutaneous injection. J Allergy Clin Immunol 2001;108:871-873.
https://doi.org/10.1067/mai.2001.119409 |
| 42 | Wortsman J, Frank S, Cryer PE: Adrenomedullary response to maximal stress in humans. Am J Med 1984;77:779-784.
https://doi.org/10.1016/0002-9343(84)90512-6 |
| 43 | Rawas-Qalaji MM, Simons FE, Simons KJ: Sublingual epinephrine tablets versus intramuscular injection of epinephrine: dose equivalence for potential treatment of anaphylaxis. J Allergy Clin Immunol 2006;117:398-403.
https://doi.org/10.1016/j.jaci.2005.12.1310 |
| 44 | Rohlich P, Anderson P, Uvnas B: Electron microscope observations on compounds 48-80-induced degranulation in rat mast cells. Evidence for sequential exocytosis of storage granules. J Cell Biol 1971;51:465-483.
https://doi.org/10.1083/jcb.51.2.465 |
| 45 | Zelechowska P, Agier J, Rozalska S, Wiktorska M, Brzezinska-Blaszczyk E: Leptin stimulates tissue rat mast cell pro-inflammatory activity and migratory response. Inflamm Res 2018;67:789-799.
https://doi.org/10.1007/s00011-018-1171-6 |
| 46 | Wasserman SI: Mast cell biology. J Allergy Clin Immunol 1990;86:590-593.
https://doi.org/10.1016/S0091-6749(05)80221-0 |
| 47 | Scanzano A, Cosentino M: Adrenergic regulation of innate immunity: a review. Front Pharmacol 2015;6:171.
https://doi.org/10.3389/fphar.2015.00171 |
| 48 | Brosnan CF, Goldmuntz EA, Cammer W, Factor SM, Bloom BR, Norton WT: Prazosin, an alpha 1-adrenergic receptor antagonist, suppresses experimental autoimmune encephalomyelitis in the Lewis rat. Proc Natl Acad Sci U S A 1985;82:5915-5919.
https://doi.org/10.1073/pnas.82.17.5915 |
| 49 | Chang TW, Shiung YY: Anti-IgE as a mast cell-stabilizing therapeutic agent. J Allergy Clin Immunol 2006;117:1203-1212; quiz 1213.
https://doi.org/10.1016/j.jaci.2006.04.005 |
| 50 | Cruse G, Duffy SM, Brightling CE, Bradding P: Functional KCa3.1 K+ channels are required for human lung mast cell migration. Thorax 2006;61:880-885.
https://doi.org/10.1136/thx.2006.060319 |
| 51 | Macrez-Lepretre N, Kalkbrenner F, Schultz G, Mironneau J: Distinct functions of Gq and G11 proteins in coupling alpha1-adrenoreceptors to Ca2+ release and Ca2+ entry in rat portal vein myocytes. J Biol Chem 1997;272:5261-5268.
https://doi.org/10.1074/jbc.272.8.5261 |
| 52 | Cotecchia S, Del Vescovo CD, Colella M, Caso S, Diviani D: The alpha1-adrenergic receptors in cardiac hypertrophy: signaling mechanisms and functional implications. Cell Signal 2015;27:1984-1993.
https://doi.org/10.1016/j.cellsig.2015.06.009 |
| 53 | Catacuzzeno L, Fioretti B, Franciolini F: Expression and Role of the Intermediate-Conductance Calcium-Activated Potassium Channel KCa3.1 in Glioblastoma. J Signal Transduct 2012;2012:421564.
https://doi.org/10.1155/2012/421564 |
| 54 | Thebault S, Roudbaraki M, Sydorenko V, Shuba Y, Lemonnier L, Slomianny C et al.: Alpha1-adrenergic receptors activate Ca(2+)-permeable cationic channels in prostate cancer epithelial cells. J Clin Invest 2003;111:1691-1701.
https://doi.org/10.1172/JCI16293 |
| 55 | Saleh SN, Albert AP, Large WA: Activation of native TRPC1/C5/C6 channels by endothelin-1 is mediated by both PIP3 and PIP2 in rabbit coronary artery myocytes. J Physiol 2009;587:5361-5375.
https://doi.org/10.1113/jphysiol.2009.180331 |
| 56 | van Zwieten PA: Pharmacology of centrally acting hypotensive drugs. Br J Clin Pharmacol 1980;10 Suppl 1:13S-20S.
https://doi.org/10.1111/j.1365-2125.1980.tb04899.x |